
限制性片段长度多态性(restriction fragment length polymorphism,RF LP )简称PCR-RFLP 分析。它主要是设计适当的扩增引物,使扩增片段包括某一一个或数个多态性的限制性内切酶识别序列,在PCR 扩增后用该限制酶切割PCR 产物,根据电泳后酶切(Amp-FLP) 对VNTR.STR 等重复序列,因重复单位数目的不同而呈现高度多态。因此利用重复序列两侧的特异性引物进行PCR 扩增,所得扩增片段具有高度多态性,这些不同长度的等位片段可用PAGE 分离。 限制性片段长度多态性(restriction fragment length polymorphism,缩写RFLP) 技术的原理是检测DNA在限制性内切酶酶切后形成的特定DNA片段的大小。因此凡是可以引起酶切位点变异的突变如点突变(新产生和去除酶切位点)和 一段DNA的重新组织(如插入和缺失造成酶切位点间的长度发生变化)等均可导致RFLP的产生。
精选百科
本文由作者推荐

限制性片段长度多态性
1980年Bostein提出的技术
中文名
限制性片段长度多态性
外文名
restriction fragment length polymorphism
作用
第一代DNA分子标记技术
提出时间
1980年
提出人
Bostein
技术简介
RFLP技术于1980年由人类遗传学家Bostein提出。它是第一代DNA分子标记技术。Donis—Keller利用此技术于1987年构建成第一张人的遗传图谱。DNA分子水平上的多态性检测技术是进行基因组研究的基础。RFLP(Restriction Fragment Length Polymorphism,限制片段长度多态性)已被广泛用于基因组遗传图谱构建、基因定位以及生物进化和分类的研究。RFLP是根据不同品种(个体)基因组的限制性内切酶的酶切位点碱基发生突变,或酶切位点之间发生了碱基的插入、缺失,导致酶切片段大小发生了变化,这种变化可以通过特定探针杂交进行检测,从而可比较不同品种(个体)的DNA水平的差异(即多态性),多个探针的比较可以确立生物的进化和分类关系。所用的探针为来源于同种或不同种基因组DNA的克隆,位于染色体的不同位点,从而可以作为一种分子标记(Mark),构建分子图谱。
当某个性状(基因)与某个(些)分子标记协同分离时,表明这个性状(基因)与分子标记连锁。分子标记与性状之间交换值的大小,即表示目标基因与分子标记之间的距离,从而可将基因定位于分子图谱上。分子标记克隆在质粒上,可以繁殖及保存。不同限制性内切酶切割基因组DNA后,所切的片段类型不一样,因此,限制性内切酶与分子标记组成不同组合进行研究。常用的限制性内切酶一般是HindⅢ,BamHⅠ,EcoRⅠ,EcoRV,XbaⅠ,而分子标记则有几个甚至上千个。分子标记越多,则所构建的图谱就越饱和。构建饱和图谱是 RFLP 研究的主要目标之一。
原理
该技术是利用限制性内切酶能识别DNA分子的特异序列,并在特定序列处切开DNA分子,即产生限制性片段的特性,对于不同种群的生物个体而言,他们的DNA序列存在差别。如果这种差别刚好发生在内切酶的酶切位点,并使内切酶识别序列变成了不能识别序列或是这种差别使本来不是内切酶识别位点的DNA序列变成了内切酶识别位点。这样就导致了用限制性内切酶酶切该DNA序列时,就会少一个或多一个酶切位点,结果产生少一个或多一个的酶切片段。这样就形成了用同一种限制性内切酶切割不同物种DNA序列时,产生不同长度大小、不同数量的限制性酶切片段。后将这些片段电泳、转膜、变性,与标记过的探针进行杂交,洗膜,即可分析其多态性结果。
分类

限制性片段长度多态性
DNA 结构在不同种类的生物体内存在着相当大的差异。随着对基因认识的不断深入,发现在同种生物的不同个体之间,尽管其蛋白质产物的结构和功能完全相同或仅存在细微的差异,但在DNA 水平却存在着差异,尤其在不编码蛋白质的区域以及没有重要调节功能的区域表现更为突出。DNA 顺序上的大多数突变是中性突变,即不影响生物体的表型,因而过去对这些突变不太重视,也无法用传统的遗传学方法来研究。但是,随着分子生物学技术的不断发展,人们从DNA 水平上直接分析生物体的突变成为可能。假如DNA 顺序中的某个碱基发生了突变,使突变所在部位的DNA 序列产生(或缺失)某种限制性内切酶的位点。这样,利用该限制性内切酶消化此DNA 时,便会产生与正常不同的限制性片段。这样,在同种生物的不同个体中会出现不同长度的限制性片段类型,即限制性片段多态性(Restriction Fragment Length Polymorphism ,RFLP )。RFLP类型
一类是由于限制性内切酶位点上发生了单个碱基突变而使这一限制性位点发生丢失或获得而产生的多态性,故称之为点多态性(point polymorphism )。这类多态性实际上是双态的,即有(+ )或无(- )。另一类是由于DNA 分子内部发生较大的顺序变化所致。这一类多态性又可以分成两类:第一类是由于DNA 顺序上发生突变如缺失、重复、插入所致。第二类是近年发现的所谓“高变区”。高变区(highly variable region ),是由多个串联重复顺序组成的,不同的个体高变区内所串联重复的拷贝数相差悬殊,因而高变区的长度变化很大,从而使高变区两侧限制性内切酶识别位点的固定位置随高变区的大小而发生相对位移。所以这一类型的RFLP 是由于高变区内串联重复顺序的拷贝数不同所产生的,其突出特征是限制性内切酶识别位点本身的碱基没有发生改变,改变的只是它在基因组中的相对位置。实际上,在DNA 顺序中,存在着大量的单个碱基的替换,但用通常所用的技术只能检测出影响到限制性内切酶识别位点上的突变。
单体型
不同的多态性切点在一特定人群中出现(+ )的频率不一样。如在一段DNA 中,切点A出现的频率为0.6 (即60 %的人含有该切点,而另外40 %的人在同一位点处不含该切点);而切点B的出现的频率为0.4 。如果这两个多态性切点(A,B)是随机相关的,那么,A、B同时存在(++ )的概率等于每个位点存在频率的乘积,即0.6 ×0.4=0.24, 即24 %的人同时含有A、B两个切点。如果它们是非随机相关的,那么同时存在的可能性将和预期的频率相差较大。这种相关的非随机性称为连锁不平衡。如果多态性切点数目为n ,则可有
2的N次方
种不同的组合,每种组合称为一种单体型(haplotype ),每种单体型随机相关的预期出现频率为各个位点频率乘积。但是,实践证明多态性切点之间并非随机相关。例如,在β珠蛋白基因簇内多态性切点2 到9 共8 个,理论应有256 种组合,即256 种单体型。但实际上只有3 种单体型,其中在希腊、意大利和亚洲印度人身上βA 染色体(携带正常β-珠蛋白基因的染色体)就占有94 %,而有些理论上的组合在实际上却是不存在的。在理论上,单体型(从切点2 到9 )的频率为0.46 ×0.48 ×0.7 ×0.27 ×0.83 ×0.52 ×0.37 ×0.32=0.0021, 而实际上该单体型的频率为0.64 ,两者相差甚大。这说明各个多态性切点之间是非随机相关的(连锁不平衡)。但也有例外,如切点10(HinfI 切点)与β基因簇各个切点都是连锁平衡的。从单体型与疾病基因的关系的研究发现,某一β-地中海贫血(简称β-地贫)基因在某一人群中与某些单体型存在着强烈的相关性,并且只有少数例外的情况。这样,在该人群中,只要我们确定了某个人的单体型就可以基本上确定该个体是否含有某种突变基因。其他
但是,即使在同一种族的人群或同一区域的人群中,这种连锁仍是不完全的。这主要表现在两个方面:第一,某一RFLP 单体型与某一突变基因相连锁,但在正常人中也能找到相同的单体型。因此,应用单体型就不能肯定地说某一个体是否存在该突变基因。但是,即使在同一种族的人群中或同一区域中的人群中,这种连锁也不相同。因此, 应用单体型就不能肯定地说某一个体是否存在该突变基因。第二、同一种突变也可以和几种单体型相关联。对于这两个情况,为了进行产前诊断,可以先通过家系调查,确定在该家系中某种单体型确与某种突变基因相关联后,再检测胎儿的单体型来确定该胎儿是还含有这种突变基因。但有时通过家系调查也无法确定某种单体型是否与某种突变相连锁,这时,就无法对其胎儿作出正确的判断。这也是RFLP 用于诊断的缺点所在。
高变区DNA与
DNA指纹人的卫星DNA 或称随体DNA 是由一些短的DNA 片段(10bp 左右)多次重复所构成的。重复片段的组成和拷贝数在不同的个体及基因组的不同位置上不一样。提取不同个体的基因组DNA 后,用其切点能识别序列为4 个碱基而又不切割该重复片段的限制性内切酶在重复片段的两侧切割基因组DNA ,然后将样品进行凝胶电泳。不同长度的重复片段(主要是由于重复单位的拷贝数不同所致)就会分开,再与含有这些重复序列的特异性探针杂交,便形成有个体特异性的放射性自显影图,亦称为DNA 指纹。
DNA 指纹的图谱

限制性片段长度多态性
取决于所用探针的核心序列(即重复序列中的重复单位)。目前所用的探针有两种,即探针33.15 。其核心序列为AGAGGTGGGCAGGTGG, 和33.6 ,即AGGGCTGGAGG 。这就是说这两种序列在人体基因组中不同的位置上分别重复不同的次数,而在不同个体的基因组中,对应位置上这两种核心序列的重复次数也不相同。这样用这两种探针之一与合适的酶切割的人体基因组DNA 片段杂交,在不同的个体将得到不同的DNA 指纹,而且探针33.5 的DNA 指纹图也不相同。DNA 指纹具有细胞稳定性和种系稳定性,是按孟德尔规律遗传的,而且杂合性高。杂合性可以这样来理解,对于由点突变引起的RFLP ,就某一个多态性切点来说只有两种多态性,即切点有(+ )或切点无(- )。而对由于高变区重复片段长度不同所引起的RFLP 来说,在基因组上某一个位置核心序列的重复次数在不同的个体不一样,比如在个体A为10 个拷贝,个体B为15 个拷贝,而个体C又可能为18 个拷贝等等。因此,在不同个体同一个相应位置上核心序列的重复次数就是多态的,而不是双态的。即使在基因组上的某一个位置处核心序列的次数一样,从而被酶切出的长度相同,但在基因组的其他位置上该核心序列重复次数又可能不同。由于DNA 指纹是按孟德尔规律遗传的,子代的DNA 指纹图可以追溯到其父母DNA 指纹图上,而在不是其父母的DNA 指纹图上则很难找到与其一样的小随体DNA 片段。在父亲的5 条可分辨的小随体DNA 片段中,有4 条处于杂合状态,即这4 条DNA 片段有不同的个体长度。因此,因高变区核心序列重复次数不同引起的RLFP 才是真正的多态性。在不同个体,这种RLFP 即DNA 指纹可以说不存在相同的。即使使用一种探针产生的DNA 指纹图无法鉴定这两个个体,如再用另一种探针便有可能将这两个个体区分开。DNA 指纹技术已被应用于亲子鉴定和法医学上对罪犯的确认等领域。限制性片段长度多态性相关的文章

三十年战争(英语:Thirty Years' War;德语:der Dreißigjährige Krieg;法语:La guerre de trente ans;西班牙语:La guerra de los treinta años;瑞典语:trettioåriga kriget;丹麦语:tredi
拿破仑·波拿巴(Napoléon Bonaparte,1769年8月15日-1821年5月5日),拿破仑一世,出生于科西嘉岛,创立法兰西第一帝国,是十九世纪法国伟大的军事家、政治家、法学家,执政期间被称为“法国人的皇帝”。
罗塞塔(英语:Rosetta),亦称拉希德(阿拉伯语:رشيد,Rashid),为埃及的海港都市。位于尼罗河三角洲西北部,距罗塞塔河口约13公里,人口约4万人。建于九世纪,是地中海地区与印度贸易的重要中继站。

星新一少年科幻安徽少年儿童出版社出版的图书《星新一少年科幻》是日本科幻之父星新一的科幻短篇精选集,各分册是《情感电视机》《梦之城》《妄想银行》《淘气的机器人》《你好,地球人》,除了收录作者各个时期的代表作品,还收录了入选语文教材和课外阅读的篇目。
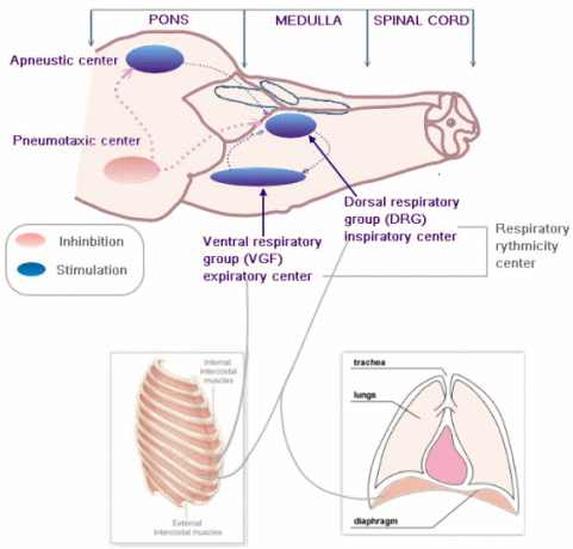
呼吸中枢参与呼吸节律产生调节的细胞群呼吸中枢(respiratory center)是指中枢神经系统内产生呼吸节律和调节呼吸运动的神经细胞群。在对呼吸中枢定位研究的诸多实验中,具有重要价值的是1923年由英国的生理学家拉姆斯登(Lumsden)对猫的脑干进行的分段横切实验。呼吸中枢分布在大脑皮层、间脑、脑桥、延髓和脊髓等各级部位,参与呼吸节律的产生和调节,共同实现机体的正常呼吸运动。延髓呼吸中枢具

尚可名片
这家伙太懒了,什么都没写!
作者

