
精选百科
本文由作者推荐
单倍群C
Y染色体DNA单倍群C
起源
单倍群C-M130似乎在M168首次发生SNP突变后不久就存在了,使单倍群CT出现,然后依次衍生出单倍群CF,单倍群C。这大概至少距今6万年前了。与 N、 O、 R、 Q、 T、 I、 J等单倍群所属的大类“F”年龄接近。
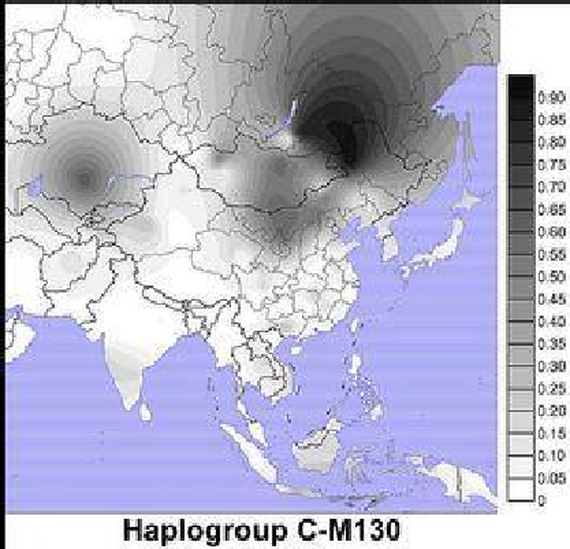
虽然C-M130在蒙古国、俄罗斯远东地区、波利尼西亚、澳大利亚等地的土著人口中最高频的存在,并且在韩国和满族人口里也中频的存在。由于现代印度人口的呈现多样性,因此猜测C-M130,要么发源,要么在印度内陆或是更广大的南亚沿海地区经受了非常久的进化。最高的多样性在东南亚被观察到,大约在4万年前从东亚地区开始向北扩张。
据信,男性携带的C-M130,距今大约6000到8000年前就已经迁移到美洲, 在北美太平洋西北沿岸的印第安纳-德内语族部落人(Na-Dene)中有着高频的分布。
亚洲也是单倍群D-M174集中的区域。然而,D-M174在关系上比C-M130更接近单倍群E,并且C-M130和D-M174的地理分布也完全是不同的,C-M130的各种亚型高频的存在于在澳大利亚土著人、波利尼西亚人、越南人、哈萨克人、蒙古人、满族人、韩国人、以及俄罗斯远东地区的土著居民中,并且在整个亚洲和大洋洲中频的分布, 其中包括了印度、斯里兰卡和东南亚。反观单倍群D,只在藏人、日本人和安达曼群岛人中才被发现,无论是在印度还是在美洲或大洋洲的土著居民中都没有发现。
单倍群C[Y染色体DNA单倍群C]
分布
C-M130通常仅分布于北亚、东亚、大洋洲和美洲人中。由于单倍群C有着非常古老的年龄,时间久远积累大量的二次突变,因此C-M130许多重要的地域性小分支已经被发现。
在欧洲殖民者到达澳大利亚之前,有高达46%的澳大利亚男性土著,不是携带C*(C-M130*),C1b2b*(C-M347*)就是携带C1b2b1(C-M210)。
男性携带C-M130低频分布的地区:印度次大陆,斯里兰卡和东南亚。另外在西班牙,来源于西班牙加泰罗尼亚格罗塔萨地区姓“Llach”的男性都携带C-M130(但来自其他地区姓该姓的男性则不携带)。在俄罗斯南部沃罗涅日的顿河沿岸出土的史前人骨遗骸中,也发现了C-M130,可追溯至34000年前,即著名的“科斯坚科14遗址”。
C在全球的分布概况如下:
C1b1a1 - M356 :印度古吉拉特邦、印度中央邦、科威特、沙特阿拉伯
C1b1a3 - Z16582:伊拉克、沙特阿拉伯
C1b1b-B68:文莱杜顺人(Dusuns)
C1b2a1a- P33:主要在大洋洲波利尼西亚人
C1b2a1d-Z32295:印尼巴朱人(Bajos)
C1b2a1c -B460:巴布亚新几内亚高伊纳贝人(Koinambes)
C1b2a3-B76:印尼巴朱人(Bajos)
C1b2b-M347:澳大利亚土著民
C2b1a1a1 - P39 :北美(土著)
C2b1a1b1 - F1756: 北亚、欧洲东部、蒙古国、中国北部等
C2b1a2 -M48:北亚、蒙古国、中国北部等
C2b1a3 - M504:北亚、中亚、蒙古国、中国北部等
C2c1a1 - CTS2657: 中国、韩国、日本、北亚、蒙古国等
C2c1a2 - Z12209: 中国、韩国、日本等
C2c1b - F845:中国、韩国、日本、越南等
C2c2 - CTS4660 : 岭南
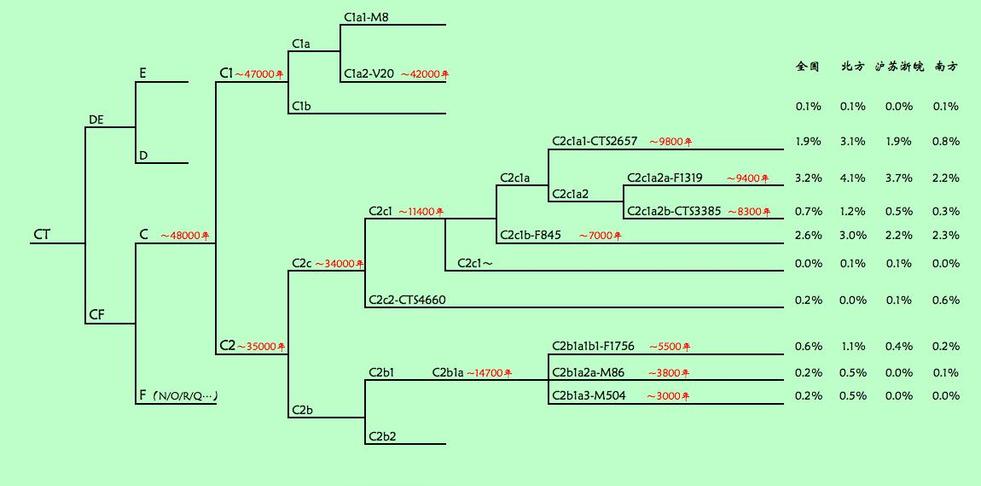
中国的C主要是C2,该分支共同祖先大约在3.5万年前。C2下面又分为南北两大支:C2b和C2c。C2b大体上集中于长城以北,在国内的满、蒙、鄂伦春等北部民族中相对集中。C2c在长城南北都有广泛分布,有些支系甚至集中于岭南(CTS4660),还有一些在南方少数民族中颇为集中(如F845下的“土司簇”)。
一些未指明的C-M130的实例,包括:
C-RPS4Y(现在称C-RPS4Y711) (xM38),这个Y-DNA在印度尼西亚的东努沙登加拉省(East Nusa Tenggara)和东帝汶共和国的人口中很常见。所有携带C-RPS4Y(xM38)的男性个体,除了亚罗群岛之外(ALOR),都被描述为操南岛语系的人。
C-RPS4Y (C-RPS4Y711) (xM38, M217),这个Y-DNA在印度尼西亚中部人群中,以相当高的频率出现。同时也在广西巴马的瑶族人口中发现(6/35),约占17%。在一份回族样本中也发现占有11%左右(4/35),在一份维吾尔族样本中发现3%(2/70),在一份赫哲族样本中发现7%(3/45),在一份鄂温克族样本中发现4%(1/26)
C-RPS4Y711(xM8, M38, M217),在一份澳大利亚原住民样本中发现48.5%(16/33),在一份瑶族样本中发现20%(12/60),在一份土家族样本中发现6.1%(3/49),在一份密克罗尼西亚人的样本中发现5.9%(1/17),在一份印尼东部人群的样本中发现5.5%(3/55),在一份印尼西部人群的样本中发现4.0%(1/25),在一份斯里兰卡人样本中3.3%(3/91),在一份马来人样本中发现3.1%(1/32),在一份印度人样本中发现2.5%(10/405),在一份巴布亚新几内亚人样本中2.2%(1/46),在一份苗族样本中1.7%(1/58),在一份维吾尔族样本中发现1.5%(1/67)。
C2
(C-M217)
单倍群C[Y染色体DNA单倍群C]
C-M217在中国北方的汉族人口中占10%左右,它的下游分支C-M401被认为可能是满族爱新觉罗氏(Manchu Aisin Gioro)的标志基因,并在中国北方的10个少数民族中都有发现,但在黑龙江、甘肃、广东、四川和新疆的汉族人口中缺失。而C-M48在现代通古斯人之中分布频率更高。东亚人以C-F1067为主,包括一个大的分支C-F2613和一个小的分支C-CTS4660。其中小的分支C-CTS4660在福建汉族和傣族中曾被发现。大的分支C-F2613在中国人(包括傣族),韩国人,日本人,越南人,孟加拉人,巴基斯坦人,哈扎拉人和车臣人中检测到,包括一些人口众多的进化分支C-F845, C-CTS2657和C-Z8440。
C-CTS2657下游有一个值得注意的分支C-M407,它在新石器时代后期扩张,在现代的布里亚特人,索约特人和哈姆尼干人中高频存在,此外还在许多卡尔梅克人和蒙古人中存在。另在300例韩国男性中样本中检测到2例,占0.67%。
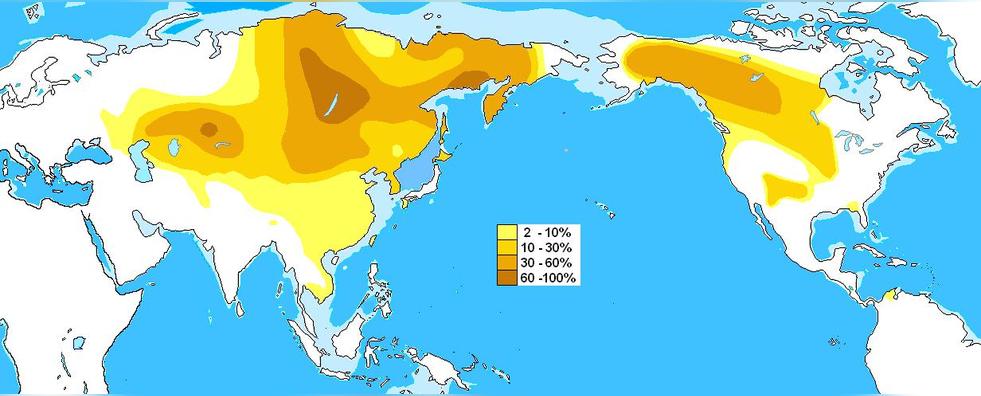
C-M217从欧洲中部和土耳其横向蔓延到南美哥伦比亚和委内瑞拉境内的印第安瓦尤人部落,并且从阿拉斯加的阿萨巴斯卡人那里纵向蔓延至越南和马来群岛。在东欧某些地区中也低频率的存在,可能是中世纪时入侵或迁移至欧洲的匈人、突厥人或蒙古人遗留下来的。
C2(M217)在布里亚特人,达斡尔人,哈扎拉人,伊捷尔缅人(堪察加人),卡尔梅克人,科里亚克人,满族人,蒙古人,鄂伦春人等人群中高频的存在。在锡伯人等其他一些通古斯语民族中,以及韩国人,阿伊努人,尼夫赫人,阿尔泰人,图瓦人,乌兹别克人,汉人,土家族人,哈尼族人和回族人中,也有适量的分布。C-M217分布最高频的地方是蒙古人群和俄罗斯远东地区。
C-M217在韩国人中的分布比以前认为的更高(16.5%)。除了哈萨克人外,C-M217在中亚地区说突厥语的民族中都是比较低频的存在,却在蒙古人和说通古斯语的民族中高频存在。C-M217可能与历史上之前的草原部落联盟一样,是13世纪蒙古人西征时 传播到突厥人中的。
然而, 在其他的日本人样本中,C-M217的分布率只有1%-3%。在2014年发表的一项研究中,对七个不同城市的日本男性进行的大样本检测,C-M217的分布率从最低的5%(15/302,札幌),到最高的7.8%(8/102,福冈),总平均率在6.1%左右(146/2390)。同时研究人员发现,C-M217或C-M8在分布频率上并没有明显的地理梯度差异。总的来说C-M217在日本的分布频率似乎与C-M8的频率是一样的,两者都占日本男性人口的5%左右。
C-M217在中国国内的几个研究的样本中,分布频率在6%-12%之间。同时在中国中部和南部的少数民族样本中也发现了同样的结果,比如湖南吉首的土家族(9/49=18.4%),哈尼族(6/34=17.6%),苗族(2/58=3.4%),畲族(1/34=2.9%),广东连南瑶族(1/35=2.9%),藏族(4/156=2.6%),以及台湾高山族(1/48=2.1%)。
在越南河内的一份样本中,C-M217占12.5%(6/48),在另一份河内京族(Kinh)的样本中占11.8% (9/76),在另一份越南平顺省占族(Cham people)的样本中8.5% (5/59)。C-M217在东南亚其它地区的分布频率较低,包括老挝人(1/25=4%,琅勃拉邦),爪哇人 (1/37=2.7%),尼泊尔人(2/77=2.6%,加德满都),泰国人(1/40=2.5%,清迈),13/500=2.6% (泰国北部),菲律宾人(1/48=2.1%,1/64=1.6%),巴厘岛(1/641=0.2%)。
虽然C-M217在西藏和尼泊尔较低频的存在(<5%),但卡西族生活的印度东北部梅加拉亚邦(Meghalaya),是一个相对高频的C-M217孤岛。卡西人(Khasi)是印度东北部的一个土著部落,他们是当地人口的主要组成部分,操卡西语或是藏缅语族(Tibeto-Burman)。根据一份2007年的调查显示,主要居住在梅加拉亚邦西部加罗丘陵(Garo Hills)地区的加罗人样本中,C-M217(xM93, P39, M86)占8.5%。而在梅加拉亚邦东部的8个卡西人部落中则占7.6%(27/353),其中Nongtrai部落最高,占(6/18=33.3%),其次是Lyngngam部落占10/60=16.7% ,War-Khasi部落占2/29=6.9%,Pnar部落占3/44 =6.8%,War-Jaintia部落占1/19=5.3%,Khynriam部落占3/87=3.4%,Maram部落占2/64=3.1%,Bhoi部落占0/32=0%。
C-M217在北美洲土著印第安纳-德内语部落人中也有着高频的分布。
C-M93 (C2a),在日本人中偶尔发现。
C-M48 (C2b1b,旧称C3c),在北通古斯人,哈萨克人,卫拉特人,卡尔梅克人,蒙古人,尤卡吉尔人,尼夫赫人,科里亚克人,伊捷尔缅人(堪察加人),乌德盖人中高频的分布。在南通古斯人,布里亚特人,图瓦人,雅库特人,楚科奇人,吉尔吉斯人,维吾尔人,乌兹别克人,卡拉卡尔帕克人和塔吉克人中有适量的分布。
C-P53.1(C2c),在新疆的锡伯族男性中,大约占10%,并且在蒙古人,鄂温克人,宁夏回族、藏族、新疆维吾尔人和甘肃汉族人口中低频的存在。
C-L1373/F1396(C2b),在中亚地区被发现。
C-P39(C2b1a1a),在北美的几个印第安土著民族中发现,包括纳-德内人,阿尔冈琴人和苏族人。
C-F2613/Z1338(C2e),在中亚和东亚都有分布。
C1
(C-F3393)
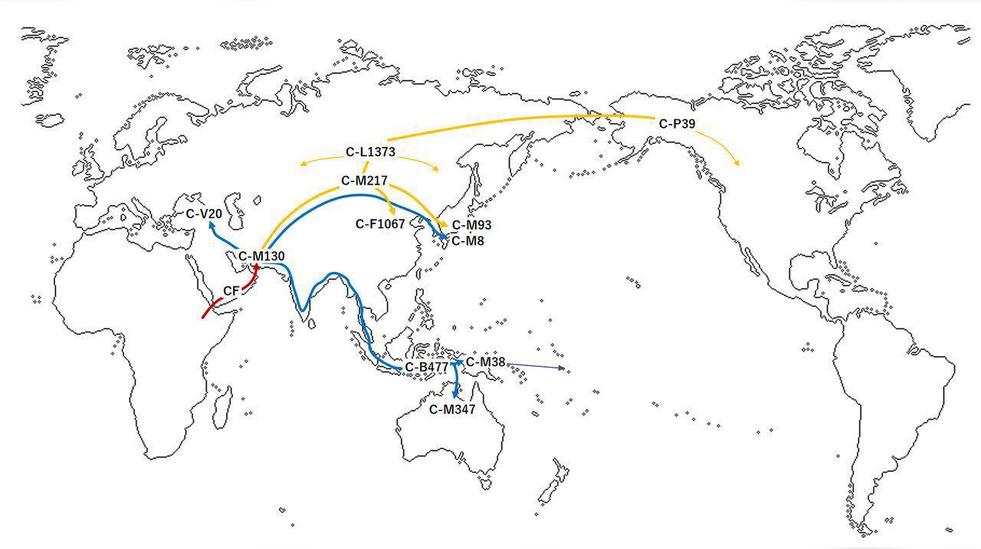
单倍群C1,也称为C-F3393,是C单倍群的两个重要分支之一。整个C单倍群分为 C1(C-F3393)和C2(C-M217),C1则分为C1a和C1b两支,C1a则又分为两支C1a1(C-M8)和C1a2。C1b在大洋洲和亚洲的部分地区很常见,反之C1a在世界范围内是极其罕见的,主要是分布在日本(C1a1)或欧洲,以及在旧石器时代的欧洲人中有发现。在尼泊尔、韩国济州岛(C1a1)、亚美尼亚人、卡拜尔人(Kabyle,生活在阿尔及利亚、突尼斯和摩洛哥等地的柏柏尔人之一) 以及中国辽宁的人群中都只发现过单个样本。
C1a下游的C1a1和C1a2两个分支在伊朗附近分开后,C1a1走北线经由阿尔泰山脉,再经过中国东北,朝鲜半岛,进入日本列岛,但C1a1在日本的阿伊努人中并没有被发现,推测由日本九州地区,向四国,冲绳方向扩散,其中一部分C1a1群体也许和日本历史上出现的隼人族有关。
C1的分支C-F3393特别是C1b,主要分布在澳大利亚的土著居民以及一些大洋洲的岛屿,少部分分布在印尼的巽他群岛(Sunda Islands)。其它的分支以非常低的频率,遍布在欧亚大陆和邻近岛屿上的那些隔绝地区。
C1a (CTS11043)
C1a* (CTS11043)还没发现任何一个样本
C1a是一种非常罕见稀少的类型,而且在地理上分布也非常的分散:
C1a1(C-M8) 在现代日本发现的频率较低 (全国平均5%)。
日本国内的比例
德岛 10% 冲绳 6.8% 本州 4.9%(青森 7.7%, 静冈4.9%) 九州 3.8%
韩国国内的比例
0.5%到1%之间
据一份祖籍朝鲜咸镜道的中国延边朝鲜族的201人样本的数据,C1a1(C-M8)的比例约1.5%(3人)。
C1a1(C-M8),作为一个古老但极其罕见的分支,其下游一分支C1a1a1,只特定出现在日本人,琉球人,以及韩国人中。另外一个分支C1a1a2,来源于中国辽宁锦州汉族人群样本,据估计这支应为东北地区土著类型。
C1a2(旧称C6)或C-V20,似乎只在欧洲、亚美尼亚、卡拜尔和尼泊尔的男性中发现。C1a2在捷克韦斯托尼采的旧石器时代晚期遗址群中也有发现。同时在俄罗斯索米尔(Sungir)一个古代猛犸象狩猎者遗址里,在两具28000年前的孩童尸体的尸体上也测到,这两具童孩身上穿着缀有上万颗猛犸象牙珠饰的衣服。
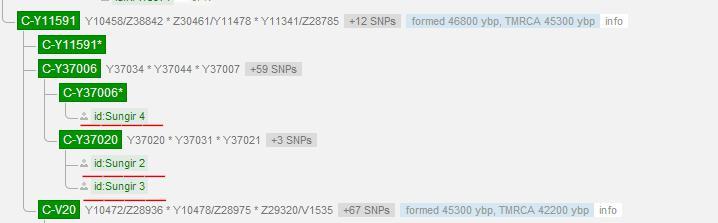
单倍群C[Y染色体DNA单倍群C]

单倍群C[Y染色体DNA单倍群C]
C-V20 (C1a2,旧称C6),在南欧国家中发现低频的分布,西班牙的拉-布拉纳(La Braña)地区,在一具距今7000年前的狩猎者遗骸中检测到。并且也同时出现在匈牙利。2016年,在比利时南部那慕尔省的戈耶山洞(Goyet Caves),在一具距今35000年前的狩猎者遗骸中,以及在捷克摩拉维亚一个叫“下韦斯托尼采(Dolni Vestonice)”的旧石器时代晚期猛犸象狩猎者营地遗址出土的一个距今30000年前的人骨中都有检测到。
C1b*(F1370),在一个名为“科斯坚科14遗址”的37000年前旧石器时代的人骨中被检测到。它也在中东地区的一小部分男性中也被发现。
C1b2(C-B477)是C-M38和C-M347的共同祖先。在欧洲人移居澳大利亚之前,它在澳大利亚的土著男性中,很有可能超过40%,属于C1b2b(C-M347)分支。在C-M347内部,至少有两个分支:C1b2b1(DYS390.1del,M210)和C1b2b1下游一个尚未识别的分支(M347xDYS390.1del,M210)。
C-P33(C1b2a1a),在波利尼西亚男性中高频的存在。
C-M356(C1b1a1,旧称C5),在印度,尼泊尔,巴基斯坦、阿富汗、阿拉伯和中国北方地区的样本中检测到低频的分布。
C-B66/Z16458 (C1b1a),在南亚,中亚和西南亚地区以较低的频率出现。
C-P55(C1b3),是在新几内亚高原地区发现的。
谱系树

单倍群C[Y染色体DNA单倍群C]
参考资料
[1]
C单倍群分化树 · C单倍群分化树[引用日期2021-11-30]
单倍群C相关的文章

华山古称太华山,位于中国陕西省渭南市华阴市城南,为中华五岳之西岳。华山南依秦岭,北临渭水,雄踞关中平原东部,以“奇险天下第一山”闻名于世,也有”华山自古一条路“的说法,景区占地面积约148平方千米。

巴士拉(البصرة,Basra),为伊拉克巴士拉省省会,位于底格里斯河和幼发拉底河交汇的夏台·阿拉伯河西岸,南距波斯湾55公里,是伊拉克第一大港及第二大城。建于635年,曾被战火摧毁,891年被重建。2003年时,全省人口估计约2,600,000人,而巴士拉城则有约1,880,000人。
迦玛尔·阿卜杜尔·纳赛尔(阿拉伯语:جمال عبد الناصر/英语:Gamal Abdel Nasser,1918年1月15日–1970年9月28日),是埃及的第二任总统,前任为纳吉布,他被认为是历史上最重要的埃及领导人之一。
拉斯维加斯(LasVegas)是美国内华达州最大的城市,也是座享有极高国际声誉的城市。拉斯维加斯建市于1905年5月15日,因位于内华达州的沙漠边缘、边境,所以拉斯维加斯全年高温。

尚可名片
这家伙太懒了,什么都没写!
作者

