
半保留复制(semiconservative replication):一种双链脱氧核糖核酸(DNA)的复制模型,其中亲代双链分离后,每条单链均作为新链合成的模板。因此,复制完成时将有两个子代DNA分子,每个分子的核苷酸序列均与亲代分子相同。子代DNA分子中,一条链来自亲代,另一条链为新合成的链。这是1953年沃森(J.D.Watson)和克里克(F.H.C.Crick)在DNA双螺旋结构基础上提出的假说,1958年得到实验证实。
精选百科
本文由作者推荐

半保留复制
双链脱氧核糖核酸的复制模型
中文名
半保留复制
外文名
semiconservative replication
提出者
(前苏联)尼古拉·科尔佐夫
提出时间
1927年
应用学科
生物学
适用领域
真核生物
简介
定义及发现过程半保留复制阐述了在所有已知细胞中DNA复制的机制。半保留复制的名字来源于这样的事实,在复制产生的两个子代DNA拷贝中,每个拷贝的DNA双链包含一个来自亲代DNA的单链和一个新合成的DNA单链。
DNA的半保留复制假说最早由前苏联生物学家尼古拉·科尔佐夫(Nikolai Koltsov)于1927年提出。1953年沃森(J.D.Watson)和克里克(F.H.C.Crick)发表的DNA双螺旋结构为此假说提供了结构上的依据。1958年美国科学家马修 梅塞森(Matthew Meselson)和富兰克林 斯塔尔(Franklin Stahl )的DNA同位素标记试验 证实了DNA的双螺旋结构和半保留复制机制。
生物学意义DNA既然是主要的遗传物质,它必须具备自我复制的能力,即通过复制形成新的和原来一样的DNA分子的能力。但双链DNA是如何解链、如何进行复制和如何保证DNA序列不变的,一直有很多的假说。
DNA在活体内的半保留复制特征已为1958年以来的大量试验所证实。DNA分子独特的双螺旋结构,为复制提供了精确的模板,通过碱基互补配对,保证了复制能够准确地进行。DNA分子通过复制,使遗传信息从亲代传给了子代,从而保持了遗传信息的连续性。DNA的这种复制方式对保持生物遗传的稳定具有非常重要的作用。
其他DNA复制方式DNA还可能存在其他两种复制方式,都以原来亲本DNA双链分子作为模板链。
全保留复制(conservative replication):保守复制会使两条原始模板DNA链以双螺旋结合在一起,并产生由两条含有所有新DNA碱基对的新链组成的拷贝。
分散复制(dispersive replication):分散复制将产生两个DNA拷贝,两个拷贝都含有由原始链或两个新链组成的不同DNA区域。
制作过程
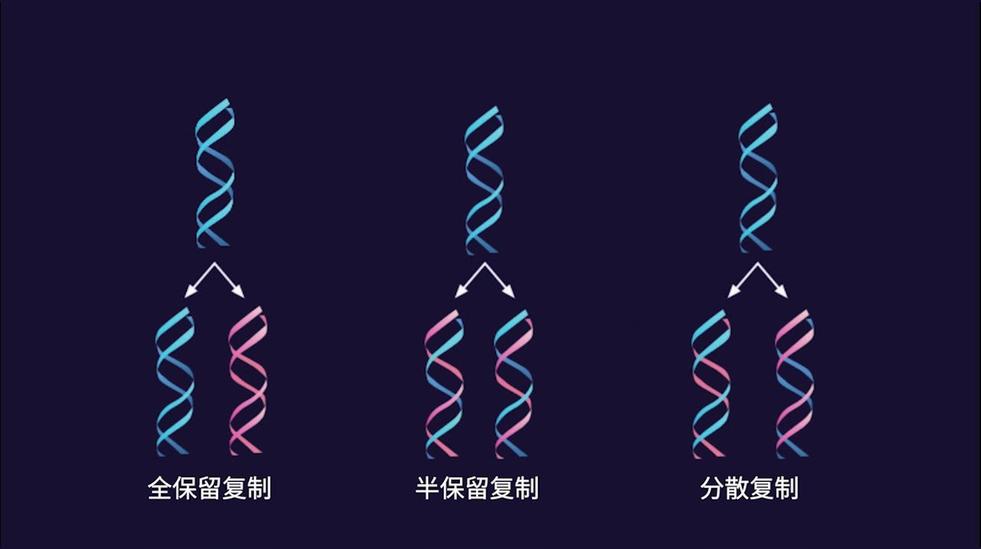
半保留复制
1958年Meselson和Stahl利用氮标记技术在大肠杆菌中首次证实了DNA的半保留复制,他们将大肠杆菌放在含有15N标记的NH4Cl培养基中繁殖了15代,使所有的大肠杆菌DNA被15N所标记,可以得到15N-DNA。然后将细菌转移到含有14N标记的NH4Cl培养基中进行培养,在培养不同代数时,收集细菌,裂介细胞,用氯化铯(CsCl)密度梯度离心法观察DNA所处的位置。由于15N-DNA的密度比普通DNA(14N-DNA)的密度大,在氯化铯密度梯度离心(density gradient centrifugation)时,两种密度不同的DNA分布在不同的区带。边解旋边复制。发生在有丝分裂间期或减数第一次分裂前的间期.遗传物质DNA既然是主要的遗传物质,它必须具备自我复制的能力。瓦特森和克里克(1953)在提出DNA双螺旋结构模型的同时,对DNA复制也进行了假设。他们根据DNA分子双螺旋结构模型,认为DNA分子的复制,首先是从它的一端氢键逐渐断开。当双螺旋的一端已拆开为两条单链时,各自可以作为模板,从细胞核内吸取与自己碱基互补的游离核苷酸(A吸取T,C吸取G),进行氢键的结合,在复杂的酶系统的作用下,逐渐连接起来,各自形成一条新的互补链,与原来模板单链互相盘旋在一起,两条分开的单链恢复为双链DNA分子,与原来的完全一样。DNA的这种复制方式称为半保留复制(semiconservative replication),因为通过复制所形成的新的DNA分子,保留原来亲本DNA双链分子的一条单链。
DNA在活体内的半保留复制特征已为1958年以来的大量试验所证实。DNA的这种复制方式对保持生物遗传的稳定具有非常重要的作用。
还可能存在其他两种复制方式,都以原来亲本DNA双链分子作为模板链。一种方法称为全保留复制(conservative replication),在复制过程中新的DNA分子单链结合在一起,形成一条新的DNA双链,而亲本DNA双链仍然被保留在一起。另一种方法称为散布式复制(dispersive replication),在复制过程中亲本DNA双链被切割成小片段,分散在新合成的两条DNA双链分子中。

半保留复制
1953年J.D.Watson和 F.H.C. Crick在提出DNA双螺旋结构时,对其互补关系予以很大的重视,而且提出了DNA的复制模型。DNA在进行复制时各以双链中的每一条链作为模板,各个和互补的前体单核苷酸配对重合而形成与这二条单链各各对应的双重子螺旋二条。所谓互补就是指腺嘌呤一定只与胸腺嘧啶配对,鸟嘌呤一定只与胞嘧啶配对,新的单核苷酸排列在模板上时,其排列法是依原来链上的碱基通过互补来决定的。这样无论子分子与子分子间,还是子分子与母分子间,碱基排列顺序是完全相同。这样一来具有和亲本完全一样的遗传信息的子分子自我增殖了二倍。这时所产生的子双重螺旋分子一条链是从亲代原封不动的接受下来的,只有相对的一条链是新合成的,所以把这种复制方式称作半保留复制。这个模型曾用重同位素标记的DNA以密度梯度离心法进行分析,或用放射性同位素标记的DNA以放射自显影法进行测定等等,用几种不同原理的方法,曾在从人到病毒的许多种生物中进行了验证,肯定了这个模型的正确性和普遍性。关于DNA是以半保留方式复制这一点已被认为是生物学中最基本的肯定性原理。DNA分子单位构成DNA分子的基本单位是脱氧核苷酸,许许多多脱氧核苷酸通过一定的化学键连接起来形成脱氧核苷酸链,每个DNA分子是由两条脱氧核苷酸链组成。DNA分子结构的特点是:①DNA分子的基本骨架是磷酸和脱氧核糖交替排列的两条主链;②两条主链是平行但反向,盘旋成的规则的双螺旋结构,一般是右手螺旋,排列于DNA分子的外侧;③两条链之间是通过碱基配对连接在一起,碱基与碱基间是通过氢键配对在一起的,其中A与T以2个氢键相配对,C与G之间以3个氢键配对。所以在一个DNA分子中,G和C的比例较高,则该DNA分子就比较稳定。
稳定性DNA分子结构具有相对的稳定性是由两个方面决定的。一是基本骨架部分的两条长链是由磷酸和脱氧核糖相间排列的顺序稳定不变;二是空间结构一般都是右旋的双螺旋结构。DNA分子的稳定性主要由三种化学键维持:互补碱基之间的氢键,碱基之间的碱基堆集力,以及主链上带负电的磷酸与溶液阳离子之间的离子键。DNA 分子的多样性是由碱基对的排列顺序的多样性决定的。DNA分子的特异性是指对于控制某一特定性状的DNA分子中的碱基排列顺序是稳定不变的,如控制合成唾液淀粉酶的基因中,不论是何人,这段DNA分子中的碱基排列顺序是稳定不变的。
结构
半保留复制
20世纪40年代~50年代,科学家已经知道DNA分子是由四种脱氧核苷酸组成的一种高分子化合物。但是,对于只由四种脱氧核苷酸组成的DNA分子为什么能够成为遗传物质,仍然
感到困惑不解。为此,许多科学家都投入到对DNA分子结构的研究
中。1953年,美国科学家沃森和英国科学家克里克,共同提出了
DNA分子的
双螺旋结构模型
(如图)。模式图DNA分子的结构模式图中可以看出,DNA分子的基本单位是脱氧
核苷酸
(如图)。由于组成脱氧核苷酸的碱基只有四种:腺嘌呤(A)、鸟嘌呤(G)、胞嘧啶(C)和胸腺嘧啶(T),因此,脱氧核苷酸也有四种,即腺嘌呤脱氧核苷酸、鸟嘌呤脱氧核苷酸、胞嘧啶脱氧核苷酸和胸腺嘧啶脱氧核苷酸。DNA分子就是由很多个脱氧核苷酸聚合而成的长链,简称多核苷酸链。结构特点沃森和克里克认为,DNA分子的立体结构是规则的双螺旋结构。这种结构的主要特点是:(1)DNA分子是由两条链组成的,这两条链按反向平行方式盘旋成双螺旋结构。(2)DNA分子中的脱氧核糖和磷酸交替连结,排列在外侧,构成基本骨架;碱基排列在内侧。(3)DNA分子两条链上的碱基通过氢键连结成碱基对,并且碱基配对有一定的规律:A(腺嘌呤)一定与T(胸腺嘧啶)配对;G(鸟嘌呤)一定与C(胞嘧啶)配对。碱基之间的这种一一对应关系,叫做碱基互补配对原则。在DNA分子的结构中,碱基之间的氢键具有固定的数目,即A与T之间以2个化学键相连(A=T),G与C之间以3个化学键相连(G≡C)。由于嘌呤分子(A、G)大于嘧啶的分子(C、T),因此,要保持DNA两条长链之间的距离不变,必定是一个嘌呤与一个嘧啶配对。根据碱基分子所占空间的大小,只有A与T配对,G与C配对,碱基对的长度才能大致相同。根据DNA分子的上述特点,沃森和克里克制作出了DNA分子的双螺旋结构模型。
DNA模型从制作的DNA双螺旋结构模型中可以看出,组成DNA分子的碱基虽然只有四种,但是,碱基对的排列顺序却是可以千变万化的。例如,在生物体内,一个最短的DNA分子也大约有4000个碱基对,这些碱基对可能的排列方式就有4种。碱基对的排列顺序就代表了遗传信息。由此可见,DNA分子是能够储存大量的遗传信息的。碱基对的排列顺序的千变万化,构成了DNA分子的多样性,而碱基对的特定的排列顺序,又构成了每一个DNA分子的特异性,这就从分子水平上说明了生物体具有多样性和特异性的原因。
DNA分子的复制
DNA分子的结构不仅使DNA分子能够储存大量的遗传信息,还使DNA分子能够传递遗传信息。遗传信息的传递是通过DNA分子的复制来完成的。DNA分子的复制是指以亲代DNA分子为模板合成子代DNA的过程。这一过程是在细胞有丝分裂的间期和减数第一次分裂的间期,随着染色体的复制而完成的。DNA的复制
半保留复制
DNA的复制是一个边解旋边复制的过程(如图)。复制开始时,DNA分子首先利用细胞提供的能量,在解旋酶的作用下,把两条螺旋的双链解开,这个过程叫做解旋。然后,以解开的每一段母链为模板,以周围环境中游离的四种脱氧核苷酸为原料,按照碱基互补配对原则,在有关酶的作用下,各自合成与母链互补的一段子链。随着解旋过程的进行,新合成的子链也不断地延伸,同时,每条子链与其对应的母链盘绕成双螺旋结构,从而各形成一个新的DNA分子。这样,复制结束后,一个DNA分子就形成了两个完全相同的DNA分子。新复制出的两个子代DNA分子,通过细胞分裂分配到子细胞中去。由于新合成的每个DNA分子中,都保留了原来DNA分子中的一条链,因此,这种复制方式叫做半保留复制
。由DNA的复制过程可以看出,DNA分子复制需要模板、原料、能量和酶等基本条件。物理实验
在上世纪中叶(1950s)JamesWatson和FrancisCrick提出了著名的DNA双螺旋以及双链间碱基配对的模型,根据这个模型,他们进一步提出了DNA复制的半保留模型(semiconservativemodel),虽然这个模型比当时并存的全保留模型(conservative模型)看起来简单易行的多,但始终缺乏有说服力的数据。最后在1957年,当时在Caltech作研究生的MatthewMeselson和作博士后的FranklinStahl设计并实现了这组著名的,证明了DNA复制半保留机理的实验。试验中,他们先将大肠杆菌细胞培养在用15NH4Cl作为唯一氮源的培养液里养很长时间(14代),使得细胞内所有的氮原子都以15N的形式存在(包括DNA分子里的氮原子)。这时再加入大大过量的14NH4Cl和各种14N的核苷酸分子,细菌从此开始摄入14N,因此所有既存的“老”DNA分子部分都应该是15N标记的,而新生的DNA则应该是未标记的。接下来他们让细胞们继续高高兴兴地生长,而自己则在在不同时间提取出DNA分子,利用CsCl密度梯度离心分离,最后得到每行从左到右由一个实验代码(这个数对我们来说没啥用),一个小图片,一个峰状谱和一个被叫做generation的数字组成。最后这个数字实际上反映了从加入氮14开始,细胞进行了多少次分裂(也就是进行了多少次复制),左边的黑色带状图案反映了该样品中DNA在离心管中的相应位置。大家可以想象一下这一列小图片是一组叠置的离心管,每根管都是管口向左比齐(在密度梯度分离法中,密度越大的分子应该越接近管底,就上图来说也就是越靠右)。这样一来不难看出随着细胞的分裂,上图中DNA的密度有所减小,从而向左迁移。而当细胞分裂了一次的时候只有一个DNA带,这就否定了所谓的全保留机理,因为根据全保留机理,DNA复制应该通过完全复制一个“老”DNA双链分子而生成一个全新的DNA双链分子,那么当一次复制结束,应该一半DNA分子是全新(双链都完全只含14N),另一半是“全老”(双链都完全只含15N)。这样一来应该在出现在离心管的不同位置,显示出两条黑带。DNA带的密度应当介于两者之间,也就是相当于一根链是14N,另一根链是15N。而经历过大约两次复制后的DNA样品(generation=1.9)在离心管中显示出强度相同的两条黑带,一条的密度和generation=1时候的一样,另一条则等同于完全是14N的DNA。这样的结果跟半保留机理推测的结果完美吻合:就这样,关于DNA复制机理的争论终于被Meselson和Stahl完美解决,而基因学和基因组学也得以在此后的五十年取得一系列重大突破。
机理证明
半保留复制
Watson与Crick在提出DNA双螺旋模型是,曾推测在DNA复制时,首先是DNA氢键断裂,双链彼此分开,然后每条链可分别作为模板,在其上按碱基互补原则配对,合成新的多核苷酸链。这样两个子代DNA的碱基顺序与亲代DNA碱基顺序完全相同。每个子代DNA中的一条链都来自亲代DNA,另一条是新合成的。这种复制称为半保留复制。实验结果表明:在全部由15N标记的培养基中得到的15N桪NA显示为一条重密度带位于离心管的管底。当转入14N标记的培养基中繁殖后第一代,得到了一条中密度带,这是15N桪NA和14N-DNA的杂交分子。第二代有中密度带及低密度带两个区带,这表明它们分别为15N14N-DNA和14N14N-DNA。随着以后在14N培养基中培养代数的增加,低密度带增强,而中密度带逐渐减弱,离心结束后,从管底到管口,CsCl溶液密度分布从高到低形成密度梯度,不同重量的DNA分子就停留在与其相当的CsCl密度处,在紫外光下可以看到DNA分子形成的区带。为了证实第一代杂交分子确实是一半15N-DNA-半14N-DNA,将这种杂交分子经加热变性,对于变性前后的DNA分别进行CsCl密度梯度离心,结果变性前的杂交分子为一条中密度带,变性后则分为两条区带,即重密度带(15N-DNA)及低密度带(14N-DNA)。它们的实验只有用半保留复制的理论才能得到圆满的解释。第一代分子含有一条亲代的链(用黑色素示),与另一条新合成的链(用白色表示)配对。在以后的连续复制过程中,原来亲代的两条链仍然保持完整,因此总有两个分子各具有一条原来亲代的链。Meslson?Stahl实验密度梯度离心后的DNA位置:左三管为对照;右三管为实验结果.实验依据
半保留复制
为验证DNA的半保留复制,科学家设计了如下实验:用含3H标记物的培养基处理蚕豆根尖细胞(2n=12),待其完成。次分裂后移入含有秋水仙素的普通培养液中,再让细胞连续分裂两次,通过放射自显影技术检验分裂期细胞染色体的放射性。据此实验分析回答:(秋水仙素作用不影响复制,但可抑制纺锤体的形成)带上3H标记的根尖细胞移入含有秋水仙素的普通培养基中,连续分裂两次,则第二次分裂前期细胞中有24条染色体。如果DNA的半保留复制假设成立,实验结果应为:(1)一条显示放射性,另一条没有放射性。(2)染色体只有一半显示放射性,而显示放射性的每条染色体只有一条染色单体显示放射性。
(一)第一代分裂后形成的每条染色体上的一个DNA分子两条链一条有放射性一条没有放射性,如果是半保留复制,以解旋以后的两条链为模板(一条有放射性一条没有放射性)在普通培养液中形成的两条子链都没有放射性,这样复制后的每条染色体都有一条染色单体具有放射性(以有放射性的那条母链为模板形成的)一条染色单体没有放射性(以没有放射性的那条链为模板形成的),所以第一次分裂中期每条染色体上都有放射性,而每条染色体上的两条染色单体一条有放射性一条没有放射性。
(二)在第一代分裂的基础上进行第二代分裂,由于第一代分裂中期的每条染色体上的染色单体一条有放射性一条没有放射性,所以着丝点分裂后形成的染色体就只有一半具有放射性了,而有放射性的染色体其中的DNA只有一条链具有放射性,如果是半保留复制,以解旋后的两条链为模板在普通培养液中形成两条没有放射性的子链,这样有放射性的染色体复制后形成的两条染色单体就只有一条染色单体显示放射性。
半保留复制相关的文章

急性脑梗死是指脑血供突然中断后导致的脑组织坏死。急性脑梗死多见于中老年人。急性脑梗死是由于供应脑部血液的动脉出现粥样硬化或血栓形成,使管腔狭窄甚至闭塞,导致局灶性急性脑供血不足而发病;也可因异常物体(固体、液体、气体)沿血液循环进入脑动脉或供应脑血液循环的颈部动脉,造成血流阻断或血流量骤减,而导致相
丘处机(1148年-1227年),或作邱处机,金代元初道士。小名丘哥,字通密,道号长春子,世称“长春真人”,登州栖霞县(今山东栖霞市)滨都里人,曾任道教全真道掌教,全真道龙门派创派祖师。丘处机与刘处玄、谭处端、马钰、王处一、郝大通和孙不二合称为“全真七子”,为全真道祖师王嚞(即王重阳)之徒。
嘉兴市,是中国浙江省地级市,位于浙江省东北部、长江三角洲杭嘉湖平原腹地,东接上海,北临苏州,南邻杭州,与宁波、绍兴隔江相望。市境介于东经120°18′-121°16′,北纬30°21′-31°02′之间,总面积3915平方公里。嘉兴市下辖南湖、秀洲2个区,嘉善、海盐2个县,平湖、海宁、桐乡3个县级市
欧洲西部(Western Europe),位于欧洲大陆西部,大西洋东岸,面积约500万平方公里,占欧洲总面积的一半左右。地理上,西欧可分为西欧、中欧、北欧、南欧四个部分。其中,狭义的西欧地理上包括英国、法国、荷兰、比利时、卢森堡、爱尔兰、摩纳哥等国家,广义的西欧还包括德国、奥地利、瑞士、列支敦士登等
生产建设兵团是指新中国成立后陆续组建的各类生产建设兵团,是中国通过军垦这种特殊体制达到巩固边防,发展经济,安置人员的目的,兵团成为党、政、军权合一,工、农、兵、学、商五位一体的半军事化组织和社会经济体系,尽管如今只有新疆生产建设兵团存在,但是在那段激情燃烧的岁月里,全国却陆续出现过12个兵团,3个农

巴比伦(Babylon)是世界著名古城遗址和人类文明的发祥地之一。它位于伊拉克首都巴格达以南90公里处,幼发拉底河右岸,建于公元前2350多年,是与古代中国、印度、埃及齐名的人类文明发祥地。巴比伦意即“神之门”,由于地处交通要冲,“神之门”不断扩展,成为幼发拉底河和底格里斯河两河流域的重镇。公元前2000年至公元前1000年曾是西亚最繁华的政治、经济以及商业和文化中心,这里还曾是古巴比伦王国和新巴比伦王国的首都。2019年7月6日,联合国教科文组织世界遗产大会上,伊拉克巴比伦古城遗址入选世界遗产名录。

尚可名片
这家伙太懒了,什么都没写!
作者
