
密码子(codon)是指信使RNA分子中每相邻的三个核苷酸编成一组,在蛋白质合成时,代表某一种氨基酸的规律。 信使RNA在细胞中能决定蛋白质分子中的氨基酸种类和排列次序。信使RNA分子中的四种核苷酸(碱基)的序列能决定蛋白质分子中的20种氨基酸的序列。而在信使RNA分子上的三个碱基能决定一个氨基酸。
精选百科
本文由作者推荐

密码子
三联体密码
中文名
密码子
外文名
genetic code
别名
三联体密码
学科
生物学
简介
mRNA(或DNA)上的三联体核苷酸残基序列
特点
tRNA的反密码子与mRNA的密码子互补
简介
密码子(codon):mRNA(或DNA)上的三联体核苷酸残基序列,该序列编码着一个特定的氨基酸,tRNA 的反密码子与mRNA的密码子互补。
起始密码子(iniation codon):指定蛋白质合成起始位点的密码子。最常见的起始密码子是甲硫氨酸或缬氨酸密码。
终止密码子(termination codon):任何tRNA分子都不能正常识别的,但可被特殊的蛋白质结合并引起新合成的肽链从翻译机器上释放的密码子。存在三个终止密码子:UAG,UAA和UGA。
应用
提高基因的异源表达可通过分析密码子使用模式,预测目的基因的最佳宿主;或者应用基因工程手段,为目的基因表达提供最优的密码子使用模式。3种不同的方式,目的都是利用密码子偏爱性来提高异源基因的表达。
翻译起始效应mRNA浓度是翻译起始速率的主要影响因素之一,密码子直接影响转录效率,决定mRNA浓度。如单子叶植物在“翻译起始区”的密码子偏性大于“翻译终止区”,暗示“翻译起始区”的密码子使用对提高蛋白质翻译的效率和精确性更为重要,因此,通过修饰编码区5′端的DNA序列,来提高蛋白质的表达水平将有望成为可能。
影响蛋白质的结构与功能基因的密码子偏性与所编码蛋白质结构域的连接区和二级结构单元的连接区有关、翻译速率在连接区会降低。马建民等通过聚类分析的方法研究发现,哺乳动物MHC基因的密码子偏爱性与所编码蛋白质的三级结构密切相关,并可通过影响mRNA不同区域的翻译速度,来改变编码蛋白质的空间构象。其研究所选取的蛋白质结构单位是蛋白指纹,它在很大程度上也是一种蛋白质功能单位,表明密码子偏性与蛋白质的功能也存在密切相关。改变密码子使用模式可目的性改变特定蛋白质的结构与功能。
基因定位功能密码子的使用模式在细胞核和细胞质遗传物质之间也存在差异,如核基因中的起始密码子只有ATG,而线粒体基因中的起始密码子为ATN;核基因中的终止密码子TGA在线粒体基因中用来编码色氨酸等。因此,可以通过比较密码子的使用模式,来进行真核生物核糖体在细胞内以及未知基因在基因组的定位。
预测进化规律类似的密码子使用模式,预示着物种相近的亲缘关系或生存环境。目 前已有研究通过比较密码子偏性的差异程度,来分析物种间的亲缘关系和进化历程。线粒体基因组具有母系遗传、分子结构简单、多态性丰富等优点,是一种重要的分子标记,研究其密码子使用偏好性,可以很好地用于确定动物类群的遗传分化和系统发生关系。
种类
密码子
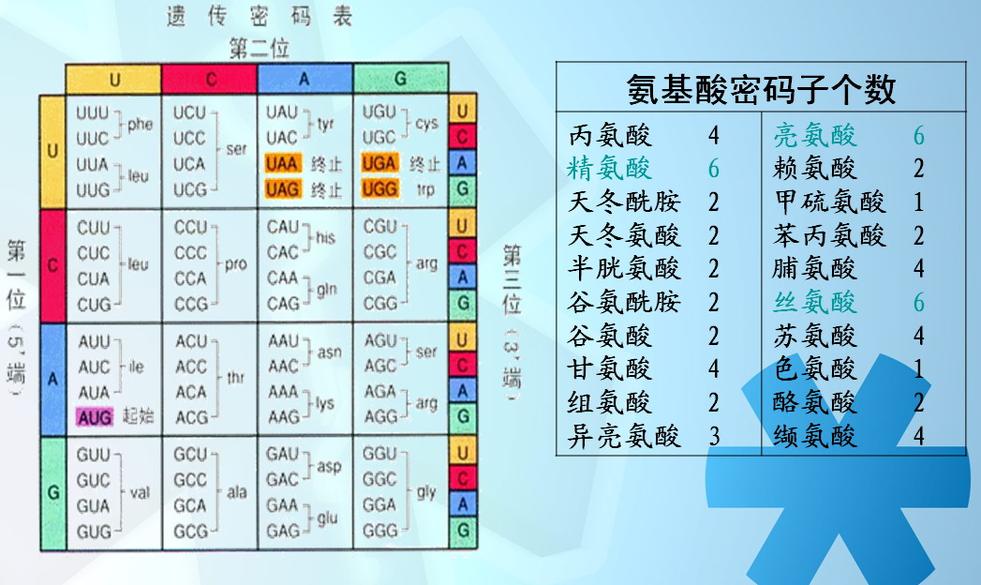
构成RNA的碱基有四种,每三个碱基决定一个氨基酸。从理论上分析碱基的组合有4的3次方=64种,64种碱基的组合即64种密码子。怎样决定20种氨基酸呢?仔细分析20种氨基酸的密码子表,就可以发现,同一种氨基酸可以由几个不同的密码子来决定,起始密码子为AUG(甲硫氨酸) ,另外还有UAA、UAG、UGA三个密码子不能决定任何氨基酸,是蛋白质合成的终止密码子。1994年版曾邦哲著《结构论》中对密码子和氨基酸的组合数学计算公式为:C1/4+2C2/4+C3/4=20氨基酸,C1/4+6(C2/4+C3/4)=64密码子。区别与联系
遗传信息是指DNA分子中基因上的脱氧核苷(碱基)排列顺序,密码子是指信使RNA上决定一个氨基酸的三个相邻碱基的排列顺序,反密码子是指转运RNA上的一端的三个碱基排列顺序。其联系是:DNA(基因)的遗传信息通过转录传递到信使RNA上,转运RNA一端携带氨基酸,另一端反密码子与信使RNA上的密码子(碱基)配对
特点
①. 遗传密码子是三联体密码:一个密码子由信使核糖核酸(mRNA)上相邻的三个碱基组成。

密码子
② 密码子具有通用性:不同的生物密码子基本相同,即共用一套密码子。③ 遗传密码子无逗号:两个密码子间没有标点符号,密码子与密码子之间没有任何不编码的核苷酸,读码必须按照一定的读码框架,从正确的起点开始,一个不漏地一直读到终止信号。
④ 遗传密码子不重叠,在多核苷酸链上任何两个相邻的密码子不共用任何核苷酸。
⑤ 密码子具有简并性:除了甲硫氨酸和色氨酸外,每一个氨基酸都至少有两个密码子。这样可以在一定程度内,使氨基酸序列不会因为某一个碱基被意外替换而导致氨基酸错误。
⑥ 密码子阅读与翻译具有一定的方向性:从5'端到3'端。
⑦有起始密码子和终止密码子,起始密码子有两种,一种是甲硫氨酸(AUG),一种是缬氨酸(GUG),而终止密码子(有3个,分别是UAA、UAG、UGA)没有相应的转运核糖核酸(tRNA)存在,只供释放因子识别来实现翻译的终止。
在信使RNA中,碱基代码A代表腺嘌呤,G代表鸟嘌呤,C代表胞嘧啶,U代表尿嘧啶(注意:RNA与DNA不同,RNA没有胸腺嘧啶T,取而代之的是尿嘧啶U,按照碱基互补配对原则,U与A形成配对)。
起源
除了少数的不同之外,地球上已知生物的遗传密码均非常接近;因此根据演化论,遗传密码应在生命历史中很早期就出现。现有的证据表明遗传密码的设定并非是随机的结果,对此有以下的可能解释:
最近一项研究显示,一些氨基酸与它们相对应的密码子有选择性的化学结合力,这显示现在复杂的蛋白质制造过程可能并非一早存在,最初的蛋白质可能是直接在核酸上形成。
尼伦伯格(M.W.Nirenberg,1927—)和马太(H.Matthaei)破译出了第一个遗传密码。
尼伦伯格和马太采用了蛋白质的体外合成技术。他们在每个试管中分别加入一种氨基酸,再加入除去了DNA和mRNA的细胞提取液,以及人工合成的RNA多聚尿嘧啶核苷酸,结果加入了苯丙氨酸的试管中出现了多聚苯丙氨酸的肽链。实验结果说明,多聚尿嘧啶核苷酸导致了多聚苯丙氨酸的合成,而多聚尿嘧啶核苷酸的碱基序列是由许多个尿嘧啶组成的(UUUUUUUU......),可见尿嘧啶的碱基序列编码由苯丙氨酸组成的肽链。结合克里克得出的3个碱基决定1个氨基酸的实验结论,与苯丙氨酸对应的密码子应该是UUU。在此后的六七年里,科学家沿着蛋白质体外合成的思路,不断地改进实验方法,破译出了全部的密码子,并编辑出了密码子表。
作用密码表

密码子
首先,这个不是生物的事实。而是基于已有的20个必需氨基酸首字母缩写,添加缺如的6个字母后得到的。依次根据氨基酸三字母缩写,中文译名拼音首字母寻找相关,再以其中密码子简并性(即重复性)最强的氨基酸为首选进行替代,具体变换为:
GCA,GCG: A→B
AGA,AGG: R→J
CCA,CCG: P→O
UUA,UUG: L→U
GUA,GUG: V→X
CAC : H→Z
对终止密码子亦进行了调整。
需要强调,这一编码方案忽略了既有的B、Z的使用,以及忽略了终止密码子间真实的强弱性。
正向翻译备选方案
A: GCU, GCC.
B: GCA, GCG.
C: UGU, UGC.
D: GAU, GAC.
E: GAA, GAG.
F: UUU, UUC.
G: GGU, GGC, GGA, GGG.
H: CAU.
I: AUU, AUC, AUA.
J: AGA, AGG.
K: AAA, AAG.
L: CUU, CUC, CUA, CUG.
M: AUG.
N: AAU, AAC.
O: CCA, CCG.
P: CCU, CCC.
Q: CAA, CAG.
R: CGU, CGC, CGA, CGG.
S: UCU, UCC, UCA, UCG, AGU, AGC.
T: ACU, ACC, ACA, ACG.
U: UUA, UUG.
V: GUU, GUC.
W: UGG.
X: GUA, GUG.
Y: UAU, UAC.
Z: CAC.
起始符:AUG【与M的编码相同,但之后需加空格□(UAA)】
空格□:UAA
断句符:UAG
终止符:UGA反向翻译:见图“修改后的密码表”
立体化学作用理论立体化学作用理论的代表人物是伍斯。他认为密码起源于氨基酸和密码子或反密码子(或更一般地和RNA)的立体化学相互作用。这个观点可以追溯至1962年,伍斯推测编码关系可能是核酸与氨基酸间的立体化学作用,他把“简并性”中涉及的密码子看作是相等的核苷酸,1965年5月,伍斯发表题为《密码的规则》的论文阐明遗传密码的排布规则,认为利用氨基酸色谱可以为分析“简并性”提供有用的证据,倾向于密码关系是一种核苷酸与氨基酸间的立体化学作用。此时,普适密码还没有完全确立,伍斯研究的编码关系还具有一定的推测性。事实上,全部密码关系的最佳立体化学匹配从来也没有被证明过。但是,氨基酸的疏水性和反密码予3’二核苷的疏水性顺序相同是已确认的事实,说明立体化学作用的因素确实重要地影响着氨基酸和反密码子的识别。
冻结偶然性理论冻结偶然性理论的代表人物是克里克。该理论认为密码关系是进化过程中的偶然性被固定下来的结果,这种关系一旦建立便永久保留下来。密码子与氨基酸的对应关系是在某个生命发生时段里被固定下来,并且很难被改变,克里克在这篇论文中讨论的编码关系来源于他在1966年冷泉港会议上呈现的密码表,这张表除了起始密码与UGA不同外,与今 天公认的普适密码是基本一致的。尽管这个假说一直受到来自密码的适应性、历史性和化学特性三方面论点的挑战,但是,从密码结构的角度,不难看出克里克当时的密码研究已经具有较强的客观性和前瞻性了。另一方面,从氨基酸的生物合成看,在一条合成路径上的几个氨基酸所用编码的密码子往往只差一个碱基。看来,后期形成的氨基酸和早期出现的氨基酸的编码存在关系,这种氨基酸和密码字典的协同进化说明编码关系并非纯属偶然。
共进化假说共进化假说提出传统的密码是从原始的简单密码进化而来,密码子的进化与氨基酸生物合成的进化是并列的。主要证据是这个原始的密码可能是由64个密码子通过高度简并只编码少量的氨基酸,而后的进化中,那些来自相关合成路径的物理化学性质不同的氨基酸却具有相似的密码子,表明密码子的进化与氨基酸生物合成具有密切相关性。王子晖认为后引入的氨基酸密码可能是通过篡夺与它具有相近生物合成路径氨基酸的密码而得到的,共进化理论鉴定了8组成对的前体和产物。这个假说后来还由朱利奥(M.Di Giulio)加以发展。
试管选择理论艾根等在研究遗传密码起源时进行试验:在试管里没有任何酶和模板的参与下,仅仅依靠锌离子的催化,将核苷酸单体聚合成寡核苷酸,并通过彼此互为模板的复制、扩增,最终在不同条件的继代培养下,优选出不同的tRNA克隆,然后形成RNA分子的准种群。这个实验被称为“试管选择性理论”,证明在无生命力作用的情况下,自然条件下完全可以形成启动生命形成的核糖核酸。根据地球形成之后的物理化学环境,推测生物大分子形成的活跃期是在距今38亿年至40亿年左右。依据实验获得的启示,研究者认为地球的早期条件会影响早期短序列RNA的产生及密码进化。但这个理论并没有对密码表结构本身给予足够的关注。
原始的遗传密码可能比今 天简单得多,随着生命演化制造出新的氨基酸再被利用而令遗传密码变得复杂。虽然不少证据证明这观点,但详细的演化过程仍在探索之中。经过自然选择,现时的遗传密码减低了突变造成的不良影响。即,遗传密码是由选择(selection)、历史(history)和化学(chemistry)三个因素在不同阶段起作用的(综合进化假说)。
氨基酸特性
氨基酸
第 一 位 碱 基 U
UUU (Phe/F)苯丙氨酸UUC (Phe/F)苯丙氨酸UUA (Leu/L)亮氨酸UUG (Leu/L)亮氨酸UCU (Ser/S)丝氨酸UCC (Ser/S)丝氨酸UCA (Ser/S)丝氨酸UCG (Ser/S)丝氨酸UAU (Tyr/Y)酪氨酸UAC (Tyr/Y)酪氨酸UAA (终止)UAG (终止)UGU (Cys/C)半胱氨酸UGC (Cys/C)半胱氨酸UGA (终止)UGG (Trp/W)色氨酸
第 一 位 碱 基 C
CUU (Leu/L)亮氨酸CUC (Leu/L)亮氨酸CUA (Leu/L)亮氨酸CUG (Leu/L)亮氨酸CCU (Pro/P)脯氨酸CCC (Pro/P)脯氨酸CCA (Pro/P)脯氨酸CCG (Pro/P)脯氨酸CAU (His/H)组氨酸CAC (His/H)组氨酸CAA (Gln/Q)谷氨酰胺CAG (Gln/Q)谷氨酰胺CGU (Arg/R)精氨酸CGC (Arg/R)精氨酸CGA (Arg/R)精氨酸CGG (Arg/R)精氨酸
第 一 位 碱 基 A
AUU (Ile/I)异亮氨酸AUC (Ile/I)异亮氨酸AUA (Ile/I)异亮氨酸AUG (Met/M)甲硫氨酸(起始)ACU (Thr/T)苏氨酸ACC (Thr/T)苏氨酸ACA (Thr/T)苏氨酸ACG (Thr/T)苏氨酸AAU (Asn/N)天冬酰胺AAC (Asn/N)天冬酰胺AAA (Lys/K)赖氨酸AAG (Lys/K)赖氨酸AGU (Ser/S)丝氨酸AGC (Ser/S)丝氨酸AGA (Arg/R)精氨酸AGG (Arg/R)精氨酸
第 一 位 碱 基 G
GUU (Val/V)缬氨酸GUC (Val/V)缬氨酸GUA (Val/V)缬氨酸GCU (Ala/A)丙氨酸GCC (Ala/A)丙氨酸GAU (Asp/D)天冬氨酸GAC (Asp/D)天冬氨酸GAA (Glu/E)谷氨酸GGU (Gly/G)甘氨酸GGC (Gly/G)甘氨酸GCA (Ala/ A)丙氨酸GCG (Ala/A)丙氨酸GUG (Val/V)缬氨酸GAG (Glu/E)谷氨酸GGG (Gly/G)甘氨酸语法及举例
语法
a.以AUG表示进入正式信息编码区;b.空格 (□) 用 UAA;c.逗号或其它文章内断续标点用 UAG;d.编码结束(最后一个句号)用 UGA表示;e. 默认为表达英语,如果需要表达其它语种,可以在起始信号(5'端)前延长6个碱基,编码语言使用语言的国家双字母缩写,如拼音加注CN,日语假名罗马体加注JP等。举例
通用语言:I love U.中转:(Start)□I□love□U.最终的核酸语言:AUG UAA AUU UAA CUG CCA GUC GAA UAA UUA UGA可以和连鹤或串珠等实体结构结合来进行空间变换,加强教学效果。但需要使游戏者谨记这套密码系统是修改过的。密码子相关的文章

小编整理:香港岛,香港特别行政区下辖行政区。香港岛(英文:HongKongIsland),简称为港岛,是中国香港最繁华的地方。香港岛简称为港岛香港岛(英文:HongKongIsland),简称为港岛,是香港最繁荣的地方。基本信息中文名香港岛外文名HongKon
1917年俄国革命是1917年于俄国发生的一系列革命运动的统称,这些运动最终灭亡了俄罗斯帝国并建立了苏维埃俄国。在1917年3月(儒略历2月)的二月革命使得罗曼诺夫王朝被推翻,临时政府掌权。同年11月(儒略历10月)的十月革命使得临时政府遭推翻,被布尔什维克(苏维埃)政府取代。
麦芽糖是一种由两个葡萄糖分子以α-1,4糖苷键连接而成的淀粉糖产品,也称为麦芽二糖。根据麦芽糖含量的不同,麦芽糖浆可分为普通麦芽糖浆、高麦芽糖浆和超高麦芽糖浆。麦芽糖是一种碳水化合物,由含淀粉酶的麦芽作用于淀粉而制得,可用作营养剂和培养基的配制。此外,麦芽糖也是一种中国传统怀旧小食。
拉贾斯坦邦是印度西部的邦,与巴基斯坦接壤。首府斋浦尔。英国统治时,拉贾斯坦邦叫拉贾普达那,意即拉吉普特人居住的地区。当时的拉贾普达那地区有18个国王和 2个萨拉达尔,因此人们又把这个地区叫做拉吉瓦拉,即“国王的地方”。

陈惠卿,台湾女性配音员,现居台湾省台北市。毕业后一直从事戏剧及广告的配音工作。曾为琼瑶剧中一些楚楚可人的女主角担任配音员。她的配音还包括公司简介、电视,电台广告、公司机关、机场、银行的语音系统等。代表作品《还珠格格》中的夏紫薇。

液氨,又称为无水氨,是一种无色液体,有强烈刺激性气味。氨作为一种重要的化工原料,为运输及储存便利,通常将气态的氨气通过加压或冷却得到液态氨。液氨易溶于水,溶于水后形成铵根离子NH4+、氢氧根离子OH-,溶液呈碱性。液氨多储于耐压钢瓶或钢槽中,且不能与乙醛、丙烯醛、硼等物质共存。液氨在工业上应用广泛,具有腐蚀性且容易挥发,所以其化学事故发生率很高。

尚可名片
这家伙太懒了,什么都没写!
作者
