碱基是指嘌呤和嘧啶的衍生物,是核酸、核苷、核苷酸的成分。DNA和RNA的主要碱基略有不同,其重要区别是:胸腺嘧啶是DNA的主要嘧啶碱,在RNA中极少见;相反,尿嘧啶是RNA的主要嘧啶碱,在DNA中则是稀有的。 除主要碱基外,核酸中也有一些含量很少的稀有碱基。稀有碱基的结构多种多样,多半是主要碱基的甲基衍生物。tRNA往往含有较多的稀有碱基,有的tRNA含有的稀有碱基达到10%。嘌呤和嘧啶碱基是近乎平面的分子,相对难溶于水:在约260纳米的紫外光区有较强的吸收。
精选百科
本文由作者推荐

碱基
嘌呤和嘧啶的衍生物
中文名
碱基
外文名
nucleobase
电子产品类别
5种
作用
组成DNA
主要成分
核酸、核苷、核苷酸
种类
腺嘌呤、鸟嘌呤、胞嘧啶、胸腺嘧啶、尿嘧啶
构成物质
ATP,GTP,CoA
性质
嘌呤和嘧啶的衍生物
构成元素
碳、氧、氢、氮
别称
核碱基、含氮碱基
原则
互补原则
定义
碱基是合成核苷、核苷酸和核酸的基本组成单位,其组成元素中含有氮,也称“含氮碱基”。
配对
在典型的双螺旋DNA中,每个碱基对都含有一个嘌呤和一个嘧啶:A与T配对通过2个氢键相连,C与G配对或Z配P或S配B是通过3个氢键相连。这些嘌呤-嘧啶间的配对现象被称为碱基互补,连接DNA两条链的碱基通常被比喻成梯子中的横档梯级。嘌呤和嘧啶间配对的部分原因是受到空间的限制,因为这种配对组合使得DNA螺旋成为一个具有恒定宽度的几何形状。A-T和C-G配对在互补碱基的胺和羰基之间形成双或三氢键。
修饰
DNA和RNA分子中还含有核酸链形成后经过修饰形成的其它非主要碱基。这些碱基大多是在上述嘌呤或嘧啶碱的不同部位甲基化(methylation)或进行其它的化学修饰而形成的衍生物。DNA中最常见的修饰碱基是5-甲基胞嘧啶(m5C)。RNA中有许多修饰的碱基,包括核苷类假尿苷(Ψ)、二氢尿苷(D)、肌苷(I)和7-甲基鸟苷(m7G)中含有的碱基。次黄嘌呤和黄嘌呤是通过诱变剂处理产生的许多修饰碱基中的两种,它们都是通过脱氨作用(用羰基取代胺基)产生的。次黄嘌呤源于腺嘌呤,黄嘌呤源于鸟嘌呤。
合成
在医学中,几种核苷类似物用作抗癌剂和抗病毒剂。病毒聚合酶将这些化合物与非主要碱基结合。病人服用的核苷类似物进入体内被转化为核苷酸而在细胞中被激活。
发现进程
生物体中常见的碱基有5种,分别是腺嘌呤(A)、鸟嘌呤(G)、胞嘧啶(C)、胸腺嘧啶(T)和尿嘧啶(U),2019年又人工合成了4种碱基,美国科学家StevenA.Benner将这4个新成员分别命名为“Z”“P”“S”“B”(顾名思义,前5种碱基中,腺嘌呤和鸟嘌呤属于嘌呤族(缩写作R),它们具有双环结构。胞嘧啶、尿嘧啶、胸腺嘧啶属于嘧啶族(Y),它们的环系是一个六元杂环。它们也被称为主要或标准碱基。它们是组成遗传密码的基本单元,其中碱基A、G、C和T存在于DNA中,而A、G、C和U存在于RNA中。值得注意的是,胸腺嘧啶比尿嘧啶多一个5位甲基,这个甲基增大了遗传的准确性。碱基通过共价键与核糖或脱氧核糖的1位碳原子相连而形成的化合物叫核苷。核苷再与磷酸结合就形成核苷酸,磷酸基接在五碳糖的第5位碳原子上)。
种类简述
新的碱基
甲基胞嘧啶(mC):源于C,是表观遗传机制的主要原因。作为一种重要的表观遗传修饰,mC参与基因表达调控、X-染色体失活、基因组印记、转座子的长期沉默和癌症的发生。
甲基腺嘌呤(mA),其主要作用是确定表观基因组的性质,并因此在细胞的生命过程中发挥重要作用。藻类、蠕虫以及苍蝇都拥有mA。mA的主要功能是调控某些基因的表达,因此,构成了一种新的表观遗传标记。
新发现的碱基还有:5-胞嘧啶甲基、5-羟甲基胞嘧啶(5-hmC)、5-胞嘧啶甲酰(5-formylcytosine)和5-胞嘧啶羧基(5-carboxylcytosine)
截至2020年已发现的碱基种类共有21种。
结构

碱基置换类型及缺失和插入突变示意图
在脱氧核糖核酸和核糖核酸中,起配对作用的部分是含氮碱基。5种碱基都是杂环化合物,氮原子位于环上或取代氨基上,其中一部分(取代氨基,以及嘌呤环的1位氮、嘧啶环的3位氮)直接参与碱基配对。碱基共有5种:胞嘧啶(缩写作C)、鸟嘌呤(G)、腺嘌呤(A)、胸腺嘧啶(T,DNA专有)和尿嘧啶(U,RNA专有)。顾名思义,5种碱基中,腺嘌呤和鸟嘌呤属于嘌呤族(缩写作R),它们具有双环结构。胞嘧啶、尿嘧啶、胸腺嘧啶属于嘧啶族(Y),它们的环系是一个六元杂环。RNA中,尿嘧啶取代了胸腺嘧啶的位置,值得注意的是,胸腺嘧啶比尿嘧啶多一个5位甲基,这个甲基增大了遗传的准确性。碱基通过共价键与核糖或脱氧核糖的1位碳原子相连而形成的化合物叫核苷。核苷再与磷酸结合就形成核苷酸,磷酸基接在五碳糖的5位碳原子上。
新成员
上世纪80年代初,由这四种“经典”DNA碱基组成的家族中迎来了第五名成员:甲基胞嘧啶(mC),其源于胞嘧啶。mC的出现引发了科学家们极大地关注,并获得了广泛的研究。上世纪90年代后期,mC被广泛看成是表观遗传机制的主要原因:它能够根据每个组织的生理需要,打开或关闭基因。而且,随着研究的进一步深入,科学家们现在知道,作为一种重要的表观遗传修饰,mC参与基因表达调控、X-染色体失活、基因组印记、转座子的长期沉默和癌症的发生。
近日,西班牙科学家在最新出版的《细胞》杂志上撰文指出,或许存在着第六种碱基——甲基腺嘌呤(mA),其主要作用是确定表观基因组的性质,并因此在细胞的生命过程中发挥重要作用。
据每日科学网4日报道,西班牙Bellvitge生物医学研究所表观遗传学和癌症生物学计划负责人、巴塞罗那大学遗传学教授曼奈·埃特雷在《细胞》杂志上发表文章,描述了第六种碱基——mA存在的可能性,他认为,这种碱基也帮助确定表观基因组,并因此在细胞生命过程中发挥着重要作用。
埃特雷在论文中表示:“早在数年前,我们就知道,在我们生物学上的远亲——细菌的基因组内就存在mA,主要作用保护其免受其他生物体遗传物质的入侵,但当时科学家们认为,这一现象只出现在原始细胞内。”
埃特雷继续解释说:“现在《细胞》杂志发表的三篇论文表明,藻类、蠕虫以及苍蝇都拥有mA,这些生物的细胞像人体细胞一样都是真核细胞,说明人体细胞内也可能拥有第六种碱基。研究表明,mA的主要功能是调控某些基因的表达,因此,构成了一种新的表观遗传标记。在我们所描述的这些基因组内,mA的浓度都很低,但随着拥有高灵敏度分析方法的发展,使得这项研究成为了可能。除此之外,mA可能也在干细胞和发育初期发挥重要作用。”
种类
近几年,有人将表观遗传学修饰——5-胞嘧啶甲基称为第5种碱基,5-羟甲基胞嘧啶(5-hmC)称为第6种碱基。在最新的研究成果中,研究人员发现了第7种,和第8种DNA碱基:5-胞嘧啶甲酰(5-formylcytosine),5-胞嘧啶羧基(5-carboxylcytosine)。这两种碱基实际上都是由胞嘧啶经由张毅教授研究组一直研究的关键蛋白:Tet蛋白修饰后形成。
作用
组成DNA
碱基
DNA(脱氧核糖核酸)的结构出奇的简单。DNA分子由两条很长的糖链结构构成骨架,通过碱基对结合在一起,就像梯子一样。整个分子环绕自身中轴形成一个双螺旋。两条链的空间是一定的,为2nm。在形成稳定螺旋结构的碱基中共有4种不同碱基。根据它们英文名称的首字母分别称之为A(ADENINE腺嘌呤)、T(THYMINE胸腺嘧啶)、C(CYTOSINE胞嘧啶)、G(GUANINE鸟嘌呤),另有U(URACIL尿嘧啶)。DNA与RNA共有的碱基是腺嘌呤、胞嘧啶和鸟嘌呤。胸腺嘧啶存在于DNA中,而尿嘧啶则存在于RNA中。每种碱基分别与另一种碱基的化学性质完全互补,嘌呤是双环,嘧啶是单环,两个嘧啶之间空间太大,而嘌呤之间空间不够。这样A总与T配对,G总与C配对。这四种化学“字母”沿DNA骨架排列。“字母”(碱基)的一种独特顺序就构成一个“词”(基因)。每个基因有几百甚至几万个碱基对。嘌呤和嘧啶都有酮-烯醇式互变异构现象,一般生理pH条件下呈酮式。AGCT(U)四种碱基在DNA中的排列遵循碱基互补配对原则
有些核酸中含有修饰碱基(或稀有碱基),这些碱基大多是在上述嘌呤或嘧啶碱的不同部位甲基化(methylation)或进行其它的化学修饰而形成的衍生物。例如有些DNA分子中含有5-甲基胞嘧啶(m5C),5-羟甲基胞嘧啶(hm5C)。某些RNA分子中含有1-甲基腺嘌呤(m1A)、2,2-二甲基鸟嘌呤(m22G)和5,6-二氢尿嘧啶(DHU)等。
构成物质碱基还构成一些生命必须物质或是重要的辅酶,如ATP,GTP,CoA等,对生命活动的作用非常大。
互补原则

碱基
(the principle of complementary base-pairing)在DNA分子结构中,由于碱基之间的氢键具有固定的数目和DNA两条链之间的距离保持不变,使得碱基配对必须遵循一定的规律,这就是Adenine(A,腺嘌呤)一定与Thymine(T,胸腺嘧啶)配对,Guanine(G,鸟嘌呤)一定与Cytosine(C,胞嘧啶)配对,反之亦然。碱基间的这种一一对应的关系叫做碱基互补配对原则。腺嘌呤与胸腺嘧啶之间有两个氢键,鸟嘌呤与胞嘧啶之间有三个氢键,即A=T,G≡C。根据碱基互补配对的原则,一条链上的A一定等于互补链上的T;一条链上的G一定等于互补链上的C,反之如此。
在DNA转录成RNA时,有两种方法根据碱基互补配对原则判断:1)将模板链根据原则得出一条链,再将得出的链中的T改为U(尿嘧啶)即可;2)将非模板链的T改为U即可。如:DNA:ATCGAATCG(将此为非模板链)TAGCTTAGC(将此为模板链)转录出的mRNA:AUCGAAUCG(可看出只是将非模板链的T改为U,所以模板链又叫无义链。这也是中心法则和碱基互补配对原则的体现。)
计算规律

5种碱基
规律一:在一个双链DNA分子中,A=T、G=C。即:A+G=T+C或A+C=T+G。也就是说,嘌呤碱基总数等于嘧啶碱基总数,各占全部碱基总数的50%。规律二:在双链DNA分子中,两个互补配对的碱基之和的比值与该DNA分子中每一单链中这一比值相等。(A1+A2+T1+T2)/(G1+G2+C1+C2)=(A1+T1)/(G1+C1)=(A2+T2)/(G2+C2)。
规律三:DNA分子一条链中,两个不互补配对的碱基之和的比值等于另一互补链中这一比值的倒数,即DNA分子一条链中的比值等于其互补链中这一比值的倒数。(A1+G1)/(T1+C1)=(T2+C2)/(A2+G2)。
规律四:在双链DNA分子中,互补的两个碱基和占全部碱基的比值等于其中任何一条单链占该碱基比例的比值,且等于其转录形成的mRNA中该种比例的比值。即双链(A+T)%或(G+C)%=任意单链(A+T)%或(G+C)%=mRNA中(A+U)%或(G+C)%。
规律五:不同生物的DNA分子中,其互补配对的碱基之和的比值(A+T)/(G+C)不同,代表了每种生物DNA分子的特异性。
相关信息

碱基配对
碱基互补配对规律的计算的生物学知识基础是基因控制蛋白质的合成。由于基因控制蛋白质的合成过程是:(1)微观领域———分子水平的复杂生理过程,学生没有感性知识为基础,学习感到非常抽象。
(2)涉及到多种碱基互补配对关系,DNA分子内部有A与T配对,C与G配对;DNA分子的模板链与生成的RNA之间有A与U配对,T与A配对,C与G配对。学习过程中,学生不易认识清楚。
(3)涉及许多数量关系(规律),在DNA双链中,①A等于T,G等于C,A+G/T+C等于A+G/T+C等1;②一条单链的A+G/T+C的值与另一条互补单链的A+G/T+C的值互为倒数。③一条单链的A+T/C+G的值,与另一条互补链的A+T/C+G的值相等;④在双链DNA及其转录的RNA之间有下列关系:一条链上的(A+T)等于另一条链上的(A+T)等于RNA分子中(A+U)等于12DNA双链中的(A+T)等,学生往往记不住。再加之转录、翻译是在不同场所进行的,学生分析问题时难以把二者联系起来。以上分析说明,关于碱基互补配对规律的计算既是教的一个难点,也是学的一个难点。教学中,如果能做到:①把复杂抽象的生理过程用简单直观的图示表现出来;②把在不同场所进行的生理过程放在一起思考;③把记忆复杂繁琐的公式(规律)转变成观察图示找出数量关系;④在计算时把表示数的符号注上脚标,以免混淆,就能轻轻松松闯过这一难关。
医学应用

碱基
DNA碱基序列决定其光敏性假设获证实。DNA分子在所有生命形态中扮演着遗传信息载体的角色,对紫外光的修改具有高度的抵抗性,但要理解其光稳定性的机制还存在一些令人费解的问题。一个重要方面是,构成DNA分子的4种碱基之间的相互作用。德国基尔大学的研究人员成功地证明,DNA链因其碱基序列而有不同的光敏感性。相关研究结果发刊登在《科学》杂志上。科学家们早就了解到,对包含在DNA中的遗传信息进行编码的个别碱基具有高度光稳定性,当它们吸收了来自紫外光辐射的能量时,这些能量会立刻再次释放。但令人惊讶的是,科学家们发现在包含有众多碱基的DNA中,这些机制变得失效或只是部分有效。因此,科学家们推断,紫外光激发的DNA分子的失活,必定由某种完全不同的、DNA特有的机制所取代。通过以各种方法测量具有不同碱基序列的DNA分子,德国的基尔大学理化研究所弗里德里希·泰姆普斯教授所领导的研究小组终于证实并阐明了该种假设。泰姆普斯教授表示,DNA通过其复杂的双螺旋结构达成其高度的光稳定性。在单股DNA链中,碱基之间的相互作用是一个堆叠在另一个之上,而且在双螺旋中,两个互补单股的碱基对之间的氢键发挥了关键作用。通过观察到的不同交互作用,DNA在某种程度上自己达成了“太阳防护”。论文作者尼娜·施瓦尔博在合成DNA分子中的过程中研究了各种不同的碱基组合。利用飞秒脉冲激光光谱学,她测量了每种组合所释放出来的特征能量。她发现,对某些碱基组合而言,这些荧光发射的“寿命”只有约100飞秒,但对其他组合而言,时间可长达数千倍。
对于该研究结果,尼娜评论道:“我们研究了光物理特性,发现不同的碱基组合具有广泛的荧光发射寿命差异,这将导致开发出一种利用激光直接识别某些遗传序列的新诊断方法,而无须像现有方法那样以染料标记DNA。”
泰姆普斯解释说,在纳米电子学领域中,合成DNA已被证明能当作“纳米线”使用。基于这些分子不同的反应时间,有朝一日或许能使用激光脉冲来“开关”特定分子。在某些情况下,甚至有可能用DNA制造出通过氢键的键合来工作的晶体管。
发展前景
涉及到RNA的试验,经常会要求对RNA分子进行固定化处理,这个过程通常由生物素进行标记,并辅以抗生物素蛋白作为支持物。人们可以将UMP、CMP之类的生物素化核苷酸单磷酸盐整合到RNA之中去,或者通过在转录反应中使用核苷酸单磷酸盐5'端衍生物类生物素,从而达到仅仅对RNA的5'端进行标注的目的。当然,人们也可以对纯化的RNA进行5'端或3'端的化学修饰。目前最简单的方法,就是在转录过程中对标记过程进行整合;但对于一些试验来说,对RNA进行特定位点的标记,比起对5'端进行标记或者为避免改变RNA的功能而仅仅使用单个标记物来说,似乎更为重要。
为达到上述目标,IchiroHirao及其在东京大学和RIKEN的合作伙伴对非天然碱基对进行了修饰,这些生物素化的碱基能被T7RNA聚合酶以特定位点的方式整合到RNA之中去。例如,2-氨基-6-(2-噻吩基)嘌呤(s)可以被整合到一个DNA模板之中去;接着,在一个标准化的转录反应中,已经被生物素化的2-氧-(1H)吡啶(y)在s补足位点被整合到了RNA转录过程中。这一方法很容易被一般性的试验室掌握,也可以通过引入T7RNA聚合酶的方式作为商业性的转录工具包加以应用。Hirao说:“除了那些包括像s和y或修饰性y底物这类非天然碱基的DNA模板外,这一工具包可在原始协议不加修改的情况下进行应用。”
在一篇新近出版的有关“核酸研究”的论文中,研究小组应用上述方法,在传感器上对一个反义的Raf-1RNA寡聚核苷酸适配子成功进行了生物素化和固定化的处理;这一寡聚核苷酸适配子准确地找到了它的目标靶点。
Hirao认为,这一由非天然碱基对组成的系统对于RNA技术将非常有用。如果这些非天然碱基对能和原核RNA聚合酶、真核RNA聚合酶一起发挥作用的话,这一系统的应用范围将大大扩展,甚至可以应用到体内试验。Hirao也计划将这一系统的应用扩展到复制、转录和翻译这些功能过程中。他说:“如果那些包含非天然碱基对的DNA片段能通过PCR手段进行扩增的话,这一系统作为工具进行使用的前景将更为广阔!”
碱基相关的文章

教育学,作为一门研究教育现象、教育问题及其规律的科学,是研究如何培养人的科学。它广泛存在于人类生活中,通过研究教育现象和教育问题,揭示教育的一般规律。教育学的任务是探讨、揭示教育规律,阐明各种教育问题,建立教育理论体系。 19世纪中叶以后,马克思主义的产生、近代心理学、生理学的发展以及现代生产和科学
亚得里亚海,意大利语作Mare Adriatico,英语作Adriatic sea。地中海的一个大海湾。长约800公里(500哩),平均宽160公里(100哩),平均水深444公尺(1,457呎),最深处1,324公尺(4,035呎),面积131,050平方公里(50,590平方哩)。北浅、南深,平
凉州,西北首府、六朝古都。凉州是今甘肃省武威市的古称,享有“天下要冲,国家藩卫”和“雍凉之都、梦幻凉州”的美称。故有《汉书》中的凉州之畜天下饶;《周书》中的凉州绯色天下最。西汉元狩二年(前121年),西汉政府为显示大汉帝国的武功军威,在西北地区原休屠王领地置武威郡,武威由此得名,新朝王莽雍州州治,东
小编整理:后凉,是十六国时期吕光建立的政权,该政权的存在时间是386年至403年。后凉政权的创建者吕光,是前秦名将。淝水之战前秦惨败后,原附属于前秦的西部地区纷纷独立,当时称为凉州的张氏也乘机自立,并上表东晋,后凉便是其中之一。淝水之战后,前秦瓦解,吕光据有凉

立原透耶作家立原透耶,有作品约30部。其作品以“轻小说”为主,著有《冥界武侠谈》系列和《CITY VICE》系列等,同时涉足奇幻、科幻、恐怖等领域。另著有《苦思乐西游传》、《迷镜楼》等短篇。其作品《卖梦的小孩子》获得1991年集英社cobalt读者大奖。其短篇小说曾经被翻译成意大利语和英语等。
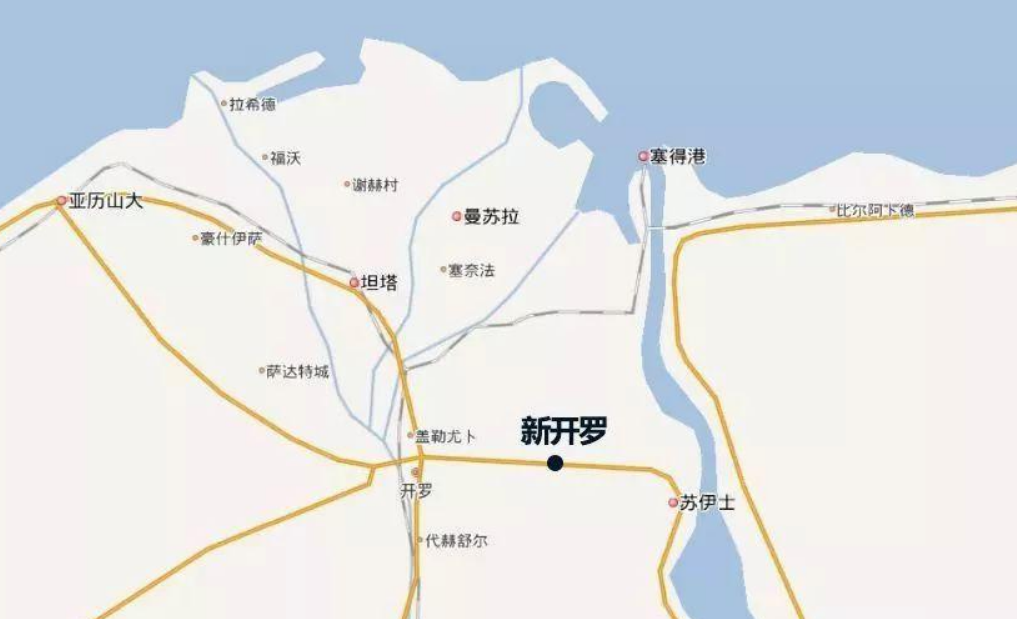
新开罗”将是一个人为创造出来的首都,可能与现在的埃及格格不入,却符合政治精英们对埃及未来的幻想。计划中的“新开罗”将是一个美丽的地方,一个“创新中心”,具有可持续发展的环境,城市居民享有高质量的生活,人们在这里可以自由呼吸,而不会因沙尘暴咳嗽不已。与之相比,“老开罗”是一个丑陋的城市。从机场到市区,一路上的建筑物都被污染物弄得脏兮兮的。耳朵里充斥着刺耳的汽车笛声,一到冬天,尼罗河上就笼罩着一层浓雾。街道上总是塞满了汽车,它们就像是这座城市的动脉血栓一样。当然,尼罗河畔会有些时尚女士穿着高跟鞋踱步,让这个城

尚可名片
这家伙太懒了,什么都没写!
作者