达尔文给本能行为所下的定义是可遗传的复杂反射,是神经系统对外界刺激所作出的先天的正确反应,这种反应已构成整个动物遗传结构的一部分。本能行为同动物的其他特征一样是通过自然选择而进化来的,是在长期进化过程中形成的。而本能行为是在"本能理论"里面,指一种先天的复杂的行为方式,它是由一组可被清晰划分的基本行为组成的:本能运动;本能运动的同义词是遗传协调。1937年康拉德·洛仑兹首次提出一种理论,其中认为这种本能行为是有所谓的钥匙刺激引起的,而且只要内在的动机一直存在,行为就会持续下去。
精选百科
本文由作者推荐
本能行为相关的文章
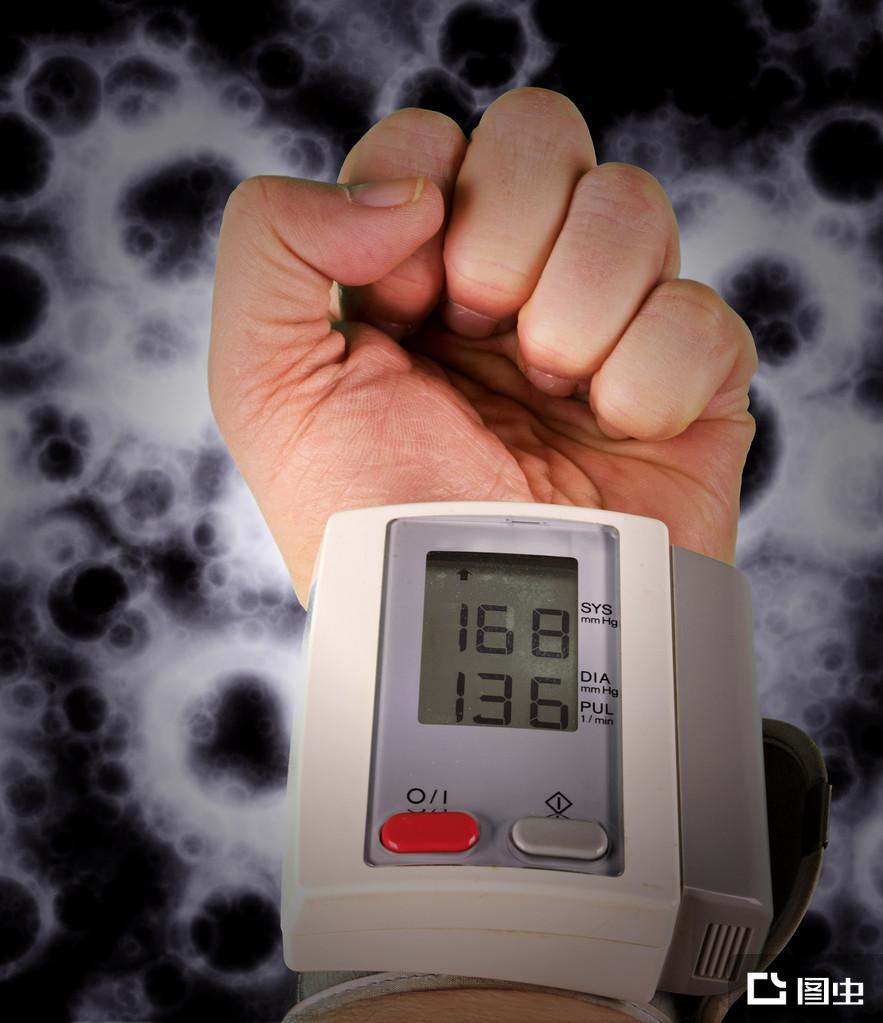
高血压(英文:Hypertension)是指在静息状态以收缩压和(或)舒张压持续升高(收缩压≥140mmHg,舒张压≥90mmHg)为主要临床表现的心血管综合征。

舟山市,是中华人民共和国浙江省下辖的地级市,位于浙江省东部、沿海地区的舟山群岛。总面积2.22万平方千米。舟山市下辖2个区,2个县,市政府驻地是舟山市新城海天大道681号。截至2021年末,舟山市常住人口116.5万人,城镇化率为72.2%。
英国皇家海军,简称为皇家海军(英语:Royal Navy,缩写:RN),是英国的海上作战力量。虽然军舰早在中世纪就被英国国王使用,并投入到百年战争期间的第一场重要海战中。但现代皇家海军起源却要追溯到16世纪初。亦是英国最古老的军种。从17世纪中叶开始,又经历18世纪先后与荷兰、法国争夺过海上霸权。
章和是东汉章帝刘炟的第三个年号,也是他的最后一个年号,共计18个月。该年号在历史上被用作几个政权的年号。据《资治通鉴》记载,元和四年七月廿七壬戌(87年9月12日),章和正式改元。在章和二年二月,汉和帝即位时沿用该年号,但次年正月初一(89年1月30日)改元永元。

杏花(学名:Prunusarmeniaca),一种花,又称杏子,是杏属李亚科植物,其果肉、果仁均可食用。杏花单生,先叶开放,花瓣白色或稍带红晕,是中国著名的观赏树木。[1]杏可配植于庭前、墙隅、道路旁、水边,也可群植、片植于山坡、水畔,是春季主要的观赏树种。杏的学名指出:西方人对杏的认识,乃源于亚美尼亚,事实上,杏广布于东亚及中亚。其可采用播种或嫁接繁殖等种植方法,一般入药的杏,就是杏的果仁,有生津、解毒、清热之效用。初夏成熟。杏树原产中国,分布很广。树龄长,可活一百年以上,既能采果又能赏花,在果木生产和

尚可名片
这家伙太懒了,什么都没写!
作者