
反义基因技术是根据核酸杂交原理设计针对特定靶序列的反义核酸, 从而抑制特定基因的表达, 包括反义RNA、反义DNA 及核酶(Ribozyme) ,它们通过人工合成和生物合成获得。反义基因技术就是从反义遗传学的角度来探索基因的结构、功能和改造等。
精选百科
本文由作者推荐
反义基因技术
反义基因技术
中文名
反义基因技术
外文名
Antisense Gene Technology
历史起源
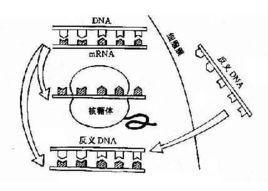
生物的遗传信息存在于多核昔酸(DNA 或RNA) 分子链上, 其碱基互补可形成双链结构。DNA不能直接表达性状或指导蛋白质的合成, 它必须通过转录mRNA并以此为模板, 合成蛋白质。在转录过程中, 双链DNA中作为模板转录mRNA的一条链称反义DNA链或称模板链。与之互补的另一条非模板链, 称有义DNA链。由有义DNA链可转录反义RNA。这种反义RNA分子在调节基因表达上有重要作用。
在分子生物学上称为干扰mRNA的互补RNA( micRNA)。现代分子生物学对于怎样产生或利用反义RNA作了深人的研究和探索, 并已取得可喜的进展。新兴发展起来的植物反义基因技术正是利用或人工合成出一段DNA(或cDNA), 再反向装上启动子和终止子, 这样反向克隆的DNA就能象正常的DNA一样, 转录出反义RNA。但这类反义RNA并不能象mRNA那样, 与核糖体结合, 翻译出蛋白质, 而是以RNA分子形式存在并与靶基因转录的RNA(包括mRNA)分子, 通过碱基互补, 形成复合体, 从而影响了RNA的剪接、加工以及与核糖体的结合, 阻止了mRNA正常的转译和表达, 导致靶基因特异性抑制或产生下向调节效果。理想的反义RNA应该与靶mRNA具特异互补性。两者在细胞核中同时存在时, 形成反义RNA-mRNA复合体, 阻止了mRNA由核内向细胞质转移, 造成特定基因失活或关闭, 产生所谓特定单基因突变现象。这在遗传学研究和植物遗传改良上很有利用价值。
反义RNA理论与技术的形成和发展是基于原核生物中天然存在的反义R N A 及其调控机理的研究而发展起来的。Tomiazwa于1981年首先报道了反义RNA的分子生物学功能, 他们在研究大肠杆菌质粒复制时, 发现反义R N A 对质粒复制有调节作用。此后在其它原核生物中也相继发现反义RNA对细菌或噬菌体的基因表达、质粒复制等起调节作用。
分类
反义基因技术主要包括:反义RNA、反义DNA 及核酶。
反义RNA根据反义RNA 的作用机制可将其分为3 类:
I类 反义RNA 直接作用于靶mRNA 的SD 序列和( 或) 部分编码区, 直接抑制翻译, 或与靶mRNA 结合形成双链RNA, 从而易被RNA 酶Ⅲ降解;
Ⅱ类 反义RNA 与mRNA 的非编码区结合, 引起mRNA 构象变化, 抑制翻译;
Ⅲ类 反义RNA 则直接抑制靶mRNA 的转录。
反义DNA反义DNA 是指一段能与特定的DNA 或RNA以碱基互补配对的方式结合, 并阻止其转录和翻译的短核酸片段, 主要指反义寡核苷酸, 因更具药用价值而倍受重视。
核酶核酶( Ribozyme) 是具有酶活性的RNA, 主要参加RNA 的加工与成熟。天然核酶可分为4 类:
I类异体催化剪切型, 如RnaseP;
Ⅱ类自体催化的剪切型, 如植物类病毒、拟病毒和卫星RNA;
Ⅲ类 第一组内含子自我剪接型, 如四膜虫人核26S rRNA;
Ⅳ类 第二组内含子自我剪接型。
作用原理
反义RNA 是通过靶RNA 进行碱基配对结合的方式参与有关的基因表达的调控。目前推测的反义RNA 作用方式有与mRNA 结合形成的二聚体阻断了核糖核蛋白体同mRNA 的结合, 从而达到了阻断翻译的目的; 与mRNA 的结合阻断了mRNA 向细胞质的运输; 与mRNA 的结合使得mRNA 易被酶识别而降解。目前尚不清楚是否还有其它的作用方式存在。
常见的获得反义RNA 的方法与基因工程方法相同。首先, 以mRNA 为模板合成互补配对的一条DNA 链, 然后以合成的互补DNA 为模板合成互补配对的另一条DNA 链, 此双链DNA 片段就是目的基因片段, 将目的基因片段反向插入适当的载体中, 然后将重组载体导入细胞, 当重组载体基因表达时, 由于是反向插入。因此, 启动子引导的不是目的基因的转录, 而是与目的基因互补配对的反义基因的转录, 从而得到反义RNA, 在实际应用中, 构建的反义基因常常只是目的基因的5′端与3′端的部分互补碱基配对序列, 但长度一般至少要大于50 bp。
植物中的应用
关于反义基因技术的研究开始也集中在原核生物, 直到真核生物自然反义系统的发现,特别是Izant 等首次证实人工构建的反义寡核昔酸在真核生物中具有生物学效应以来, 反义技术在真核生物中的研究才得以迅速发展。随其研究而发展形成的反义技术提供了一种更为直接有效的人为控制基因表达的方法而倍受生物学界关注。目前在植物领域的应用尤为凸显。
调控果实成熟
利用DNA 重组技术, 人工构建反义基因与反义RNA 系统, 调控生物体内特定基因的表达进而延长植物果实贮藏保鲜, 已成为果蔬贮藏保鲜研究工作中最活跃的领域之一。
改良作物品质
随着生活水平的提高和科学技术的进步, 人们对稻米等粮食作物食用品质的要求也越来越高。利用反义RNA 技术就可以通过对waxy 基因的遗传操作降低水稻生物体内特定基因表达水平, 来控制水稻中直链淀粉的合成, 从而改变其在胚乳中的相对含量, 达到改良稻米淀粉品质和食用品质的目的, 培育优质水稻新品种。
获得作物
雄性不育系雄性不育系( 母本) 对提高作物杂交育种效率和效果有非常重要的作用; 是作物杂交育种的技术关键, 常规的方法很难得到。目前, 用反义RNA 技术来获得雄性不育的转基因植株, 或恢复植株的雄性育性的研究工作在不同植物中取得了积极的成果, 对在杂交种子生产中的应用提供了重要的技术支持。
改变植物花色
植物的花色在园艺和商业上有重要价值, 改变花色一直是园艺学家们孜孜以求的目标。研究表明许多植物花的颜色与类黄酮有关, 苯基苯乙烯酮合成酶( CHS) 是类黄酮生物合成中起关键作用的酶;这为利用基因工程调控花色提供了理论依据。
增强
植物抗病性反义技术的建立扩展了机体抵御外来微生物的经典免疫学概念, 利用反义RNA 技术来阻断细菌、病毒在细胞中的复制, 20 世纪80 年代末期开始应用于植物抗病的研究。其原理概括就是选择细菌、病毒复制过程中的关键基因作为靶基因, 利用反义技术将其反义基因或片断转化进入植物细胞,并使其转录的RNA 结合靶基因的mRNA, 使靶基因不能正常表达, 从而达到干扰抑制细菌、病毒复制, 提高植物抗病能力的目的。经过不断完善, 反义技术在植物抗细菌、病毒的研究中得到了广泛地利用, 己经取得了积极的效果。
油料作物种子中脂肪酸合成的控制
在改变油料作物油脂的组成方面,近10年来已取得一系列重大突破, 其主要目的是增加饱和脂肪酸的含量或减少饱和脂肪酸的含量。利用反义基因技术将反义丙酮酸羧化酶(PEP基因)导入油菜, Anti-PEP转化株含油量比对照明显提高,最高的几株均增加了15%以上,并且验证了蛋白质含量与油脂含量呈显著负相关。
反义基因技术相关的文章

多疑是指神经过敏、疑神疑鬼的消极心态,属于精神心理科疾病。与生活环境、消极暗示、受过伤害等有关,因心理脆弱而过度自我保护。多疑分为内应多疑和外应多疑。内应多疑的主要表现为总是怀疑某人对自己不好,某件事对自己不利,便耿耿于怀,闷闷不乐,情绪立即反常,半天都不能排解;外应多疑的主要表现为的觉得被人怀疑后
KAA084是一个全新的折叠运动自行车设计。它作为一个紧凑型公路自行车。它的快速,硬朗,就像骑一个传统的大自行车,但它的便携能力令人难以置信。BioLogic II 框架提供了一个强大的刚性和稳定性,是其他大多数折叠车所无法比拟的水平。。一辆接近完美的小车,有着精致的外观和强大的战斗力。市内通勤,周

苏逸平《穿梭时空三千年》的作者苏逸平是继香港科幻小说家卫斯理之后,又一在港澳台及东西亚风靡至今的科幻小说家。他是生长于台湾的美籍华人,在美国学的是机电专业,但极度热爱星际间的幻想世界,坚信在冷硬的科技中,存在着复杂、柔美的人性情怀,爱恨纠葛。由他创作的“时空传奇”系列作品,有的被改编成漫画书,有的已

尚可名片
这家伙太懒了,什么都没写!
作者

