
精选百科
本文由作者推荐

基因沉寂
基因沉寂
术语
定义RNAi与转录后基因沉默(post-transcriptional gene silencing and transgene silencing)在分子层次上被证实是同一种现象。
原理基因沉寂需要经历不同的反应过程才能实现,包括组蛋白N端结构域的赖氨酸残基的去乙酰基化加工、甲基化修饰(由甲基转移酶催化,修饰可以是一价、二价和三价甲基化修饰,后者又被称为'过度’甲基化修饰(Hypermethylation) ) 、以及和甲基化修饰的组蛋白结合的蛋白质(MBP)形成“异染色质”,在上述过程中,除了部分组蛋白的N端尾部结构域需要去乙酰化、甲基化修饰之外,有时也需要在其他的组蛋白N端尾部结构域的赖氨酸或精氨酸残基上相应地进行乙酰化修饰,尽管各种修饰的最终结果会导致相应区段的基因“沉寂”失去转录活性。
作用这个“原则”就是目前尚没有真正完全清楚的“组蛋白密码”(Histone Code)。能够与甲基化组蛋白结合的蛋白质有sir1/2/3/4,这是一组被称为"Silencing Informative Repressor"的蛋白,其中,Sir2就是上文中的“去乙酰化”酶,而Sir1/3/4则负责与甲基化修饰的组蛋白结合"沉寂”相应的染色质为异染色质。此外,基因沉寂也和DNA的甲基化修饰有关,比如在真核生物基因组中的许多基因的5‘端分布有长约1KB( 千碱基对)的“CpG"岛序列(CpG island),其中的“C"芳香环5位可被甲基化修饰,之后,与甲基化修饰的DNA结合蛋白形成“沉寂"区段,使其下游基因不能表达;另外,非编码的RNA分子(non-coding RNA)也参与“基因沉默”过程。这一类型常见于含有重复DNA序列的染色质区,如着丝粒部位的基因沉寂就需要非编码RNA分子的参与。简言之,基因沉寂或者基因沉默是涉及组蛋白甲基化、去乙酰化、乙酰化,DNA的甲基化修饰,甲基化修饰组蛋白结合蛋白Sir2/3/4,甲基化DNA结合蛋白,非编码RNA等等在内的一系列复杂组分的生理反应过程。基因沉寂导致相应区段内的遗传信息不能被转录。
详细内容
动植物
基因沉默现象首先在转基因植物中发现,接着和线虫、真菌、昆虫、原生动物以及才鼠中陆续发现。大量的研究表明,环境因子、发育因子、DNA修饰、组蛋白乙酰化程度、基因拷贝数、位置效应、生物的保护性限制修饰以及基因的过度转录等都与基因沉默有关。但总的看来,基因沉默发生在两种水平上,一种是由于DNA甲基化、异染色质化以及位置效应等引起的转录水平上的基因沉默(tran-scriptional gene silencing,TGS),另一种是转录后基因沉默(post-transcriptional gene silen-cing,PTGS),即在基因转录后的水平上通过对靶标RNA进行特异性降解而使基因失活。在这两种水平上引起的基因沉默都与基因的同源性有关,称为同源依赖性的基因沉默(homology-dependent gene silencing,HDGS)。
PTGS
PTGS在多种生物中有共性,对PTGS的激活和与其相关的RNA降解调控过程有了初步的认识。也发现植物病毒在转基因植物和非转基因植物中都能和转基因一样诱发转录后基因沉默。令人吃惊的是,转基因植物的共抑制现象(转基因与同源的内源基因一起失活)、转基因植物的病毒抗性和非转基因植物对病毒正常自然侵染的抗性、真菌的quelling现象(真菌中的共抑制)、各种动物的RNA干扰(RNA interference,RNAi)以及转座因子的转座失活等这些表面看来完全不相关的现象中竟然存在着非常相似的基因沉默机制,即PTGS。这种基因沉默可能是生物体的本能的反应,因为无论是转基因、转座因子还是病毒,对植物而言都是诱发突变的外来侵入的核酸,植物为保护自己,在长期的生物进化中,形成了基因沉默这种限制外源核酸入侵的防卫保护机制。在线虫Caenorhabditis elegans中,RNAi敏感性缺失突变体中转座子的转座活性增强,表明转座子的转座失活是被一种类似PTGS的过程调控的,这一过程与RNAi作用有关,是通过细胞内双链RNA互作引起同源特异性的RNA降解。
PTGS中的RNA在细胞质中的特异性降解并不需要RNA结合到核糖体上,这与通常的RNA降解代谢调控需要与翻译相关连不同。Bass对RNAi激发的RNA特异性降解机制进了体外研究,发现通过添加外源的双链RNA或靶标mRNA可以激活PTGS,这与早期在植物中发现的双链RNA介导PTGS一致。通过对发生PTGS的转基因植物进行嫁接实验和分析植物病毒病的恢复现象观察到,PTGS是一种系统性的过程,称为系统获得性沉默(systemic acquired silencing,SAS)。PTGS的系统传播性在真菌和线虫中也得到了证明。进一步研究发现,转基因或病毒侵染介导的PTGS植物中普遍存在着大量的序列特异性正义和反义的大约25个核苷酸的小分子RNA,而正常的非转基因植物和没有发生PTGS的植物中则没有。这些小分子RNA作为信号分子,在植物中与特定的运输蛋白特异结合,防止被核酸酶的降解,通过胞间连丝和韧皮部筛管运送到植物体的各个部位,使PTGS具有系统持久性。这一运转过程与植物病毒在植物体内的运输有着十分相近的机制。这些-25nt RNA的积累需要转基因的转录或病毒的复制,与其它双链RNA等多种异常RNA相比,-25nt RNA对于PTGS的激发、靶标RNA的特异性降解以及PTGS的系统性维持更为重要。
病毒影响
早在20世纪70年代初人们就发现,病毒或类病毒侵入植物后,RNA依赖性的RNA聚合酶(RNA-dependent RNA polymerase,RdRP)的活性明显提高。RdRP是生物体内普遍存在的一种RNA聚合酶,在体外能以单链RNA或单链DNA甚至以双链RNA为模板,合成与模板互补RNA,合成的cRNA可长达100个核苷酸。1993年,Lindbo等认为在PTGS植物中的RdRP与PTGS同源依赖性的RNA特异性降解有关。最近Mour-rain等从缺失转基因介导的PTGS拟南芥突变体中分离的sgs2(suppressor of gene silencing)和sde1(silencing defective)两个基因与番茄中的RdRP基因十分相似,支持了Lindbo的观点。粗糙脉孢菌Neurospora crassa的qde-2(quilling-defective gene)和线虫C.elegans的rde-1(RNA interference-deficient gene)与蕃茄的RdRP基因也具有很高的同源性。RdRP基因剔除实验证明,RdRP对RNAi和PTGS尤为重要。这些研究结果表明,从比较低等的生物藻类、真菌到高等的植物再到动物线虫、锥虫、果蝇等中可能存在着一个由共同祖先进化来的相似的抵抗外来DNA侵入自身基因组的本能防卫机制。
RNAi引发的PTGS作为生物体中一种不完全的原始的生物免疫系统,在植物抗病毒中研究得比较详细。研究发现,植物病毒的RNA可以直接作为PTGS的激发子,并且可能通过病毒来源的基因引发转基因植物对病毒的终生系统抗性,这和PTGS的从病毒侵染点传播到整个植株以及线虫中PTGS的系统性一起证明了PTGS具有系统传播性。缺少高效PTGS的植物突变体虽然在表型上几乎没有变化。拟南芥PTGS突变体sgs2和sgs3对黄瓜花叶病毒(cucumber mosaic virus,CMV)的敏感性提高了,而突变体sde1对烟草花叶病毒(tobacco mosaic virus,TMV)和烟草脆裂病毒(tobacco rattle virus,TRV)的敏感性无变化。Baulcombe研究组认为RdRP(SDE1或SGS2)对于引发产生PTGS的双链RNA是必需的,有些RNA病毒在复制中用自身的RdRP合成双链RNA直接进入PTGS网络,而不需要寄主的RdRP去激活PTGS,这些病毒如TMV、TRV能在PTGS突变体中和野生型中一样致病,而另一些病毒如CMV需要寄主的RdRP来激活PTGS,所以PTGS突变体对这些病毒的敏感性要比野生型的高。另一方面,还可能与有些病毒产生的抑制PTGS的蛋白质有关,如CMV和番茄不孕病毒(tomato aspermy virus,TAV)的2b蛋白以及马铃薯Y病毒(potato Y virus,PVY)的蚜传辅助因(helpercomponent/protease,HC-Pro)等。由此可见,不同的RNA病毒是通过不同的位点进入并引发PTGS网络的。由于拟芥南PTGS突变体是通过转基因介导的PTGS筛选的,突变体对不同病毒敏感性的变化,也可能表明转基因和病毒侵染引发植物PTGS的机制是有差别的。在突变体sde1中,与PTGS有关的-25nt转基因特异性的RNA的积累明显减少,而病毒特异性的-25nt RNA的量不变,这也表明转基因和侵染病毒是通过不同途径引发PTGS的。
推测从大量的研究结果中我们可以推测,生物体内有一套RNA监视系统,可以通过多种异常RNA来激发。如果外来核酸是DNA(包括转基因、重组基因、DNA病毒、扩增子等),靶标RNA需要在细胞核中完全成转录后运转到细胞质中,而侵入细胞质的病毒RNA可以直接提供靶标RNA。各种不同的靶标RNA(包括与外源基因同源的内源基因和外来的DNA产生的RNA以及病毒的RNA)由寄主的RdRP或病毒自身的RdRP通过多种不同的途径反靶标RNA转变成为双链RNA,从而通过RNAi引发的PTGS。PTGS被引发后就不再需要RdRP。关于双链RNA介导的RNAi特异性靶标RNA的降解,Bass提出了这样一个假说:认为生物体内存在着一种复合酶:RNAi核酸酶,该酶具有双链RNA结合、RNase和RNA解旋酶三个活性区。首先双链RNA结合到该酶的双链RNA结合区并引导该酶识别靶标RNA,接着该酶的解旋酶完成ATP依赖性的靶标RNA与该酶结合的双链RNA的正义链的换位,RNase在靶标RNA结合位点附近完成切割,从而使靶标RNA能被进一步降解,产生大量的小片段RNA,包括序列特异性的-25nt RNA。载有序列特异性的双链RNA的游离复合酶再去识别并降解其它的靶标RNA,产生更多-25nt RNA,从而使PTGS具有持久性系统性。
结论总之,基因沉默是基因表达调控的一种重要方式,是生物体在基因调控水平上的一种自我保护机制,在外源DNA侵入、病毒侵染和DNA转座、重排中有普遍性。对基因沉默进行深入研究,可帮助人们进一步揭示生物体基因遗传表达调控的本质,在基因克服基因沉默现象,从而使外源基因能更好的按照人们的需要进行有效表达;利用基因沉默在基因治疗中有效抑制有害基因的表达,达到治疗疾病的目的,所以研究基因沉默具有极其重要的理论和实践意义。
特殊基因
与基因沉默相关特殊基因
奥地利格雷戈尔·门德尔植物分子生物学研究所日前宣布,一个包括该研究所、中国同济大学、美国加利福尼亚大学等机构科学家在内的国际科研小组发现了一种特殊基因,没有它,植物细胞内其他一些基因就只能保持沉默。
最新一期英国《自然》杂志网络版发表了这个国际科研小组的论文。这一科研小组发现的特殊基因名为RDM1,它可以编码生成一种小蛋白,从而参与指导其他基因的表达。
科学家指出,基因一般处于被保护状态中,只有通过所谓的甲基化,即与甲基接触,才能表达并发挥作用。如果去除RDM1,被保护的基因就无法甲基化,也就无法进行表达。
由于每一个植物细胞中都存在着完整的遗传信息,因此必须让某些基因保持“沉默”,植物具体的器官才能顺利地发挥各自作用。否则,所有基因就会都来表达,植物器官也将不知道听从谁的“指令”.一般在一个植物的上万个基因里,只有很少的一部分能够表达,RNA(核糖核酸)会对需要表达的基因进行标记。RDM1基因的任务就是让RNA标记过的基因表达。缺少了RDM1,植物中许多本该表达的基因就会保持“沉默”,植物无法正常生长。
两者关系
RNA剪接和基因沉默之间的联系
为了识别在RNA干涉(RNAi)和微RNA介导的基因表达调控中所涉及的因素,Gary Ruvkun及其同事对86种真核生物进行了系统发生分析,所得到的候选物再用转录和蛋白组相互作用数据进行Bayesian分析,来估计它们参与小RNA调控的概率。所识别出的小RNA辅因子中大约一半是RNAi沉默所必需的,其他当中的很多参与剪接,说明在这两个过程之间存在一个联系。
实现工具
Sigma-Aldrich公司近日在全球推出MISSION® esiRNA,一种基因特异的siRNA库,它为哺乳动物细胞中的RNAi筛选提供了一种强有力的方法。与传统的合成siRNA不同,esiRNA集合了数百个针对单个基因靶点的siRNA。这种新工具能避免鉴定有效siRNA的反复试验过程,迅速实现基因沉默,并确保脱靶效应最低。
此项技术与传统的单个siRNA介导的基因knockdown相比,优势相当明显。以前,我们需要反复试验,来鉴定出最有效的siRNA,现在有了这个siRNA的“超级库”,这一步都省了。此外,esiRNA还能确保最低的脱靶效应。esiRNA是由德国马普学会细胞生物学和遗传学(MPI-CBG)开发的。
MPI-CBG的课题组组长Frank Buchholz开创了esiRNA技术在RNAi筛选中的应用。他表示:“我们很高兴能与Sigma-Aldrich合作,让整个学术界都能享有esiRNA技术。10年前,esiRNA在美国旧金山加州大学的Mike Bishop实验室中发明。自此,它就成为我的研究重点。Sigma-Aldrich在RNAi方面的丰富经验和产品线是我们生产高效特异esiRNA的完美补充。”
哈佛大学医学院正在运用esiRNA来协助鉴定促使HIV增殖的人类基因。临床研究员Abraham Brass博士认为MISSION esiRNA的初步试验结果表明多个基因都能被非常有效地knockdown。哈佛医学院很快将会把此项技术整合到多项研究中。
esiRNA是由核酸内切酶制备的。首先用特异的引物扩增最佳的靶区域。序列经过验证后,利用RNA聚合酶产生300-600 bp基因特异的双链RNA,随后用核酸内切酶切割产生短的dsRNA。这些类似siRNA的分子经过纯化,就形成了siRNA库,它们靶定了单个基因的不同序列。
MISSION esiRNA具有下列优势:
针对每个基因靶点的MISSION esiRNA超级库
快速经济地完成初次筛选
有效的基因沉默
esiRNA的knockdown效率高于70%
脱靶效应更低
大型RNAi筛选研究的费用更省
目前,MISSION esiRNA有以下几种规格,满足不同实验的需求:
MISSION esiRNA人基因组规模库,覆盖12000多个基因靶点
MISSION esiRNA人激酶库,覆盖500多个基因靶点
定制MISSION esiRNA库(20 ug),适于10到1000个基因靶点
单个MISSION esiRNA(20 ug或50 ug),适于1到10个基因靶点
基因沉寂相关的文章

印度河是巴基斯坦主要河流,也是巴基斯坦重要的农业灌溉水源。它的名字源自梵文Sindhu(信度)之拉丁语式拼法Indus,意为“河流”。在1947年印巴分治之前,印度河是该地区的文化和商业中心地带,仅次于恒河。印度河的总长度约为2900至3200公里。印度河文明是世界上最早进入农业文明和定居社会的主要
伊玛目清真寺又称“马斯吉迪·贾米伊·阿巴斯”或“国王清真寺”,位于伊朗伊斯法罕。清真寺里外均由精美的瓷砖镶嵌而成,其外形为壁竞式的正门面向正北。清真寺的大门是镀银的,正门高达30米,门上写有许多诗文,由当时著名书法家用美丽的波斯文纳斯塔利克体书写。
武威郡,古称凉州、雍州、雍凉、姑臧、休屠、西州,西北首府、六朝古都。意为彰显大汉帝国的武功军威到达祁连山的雍凉之地,西汉元狩二年(前121年)置,今甘肃省武威市,隶属凉州刺史部,治所姑臧(今甘肃省武威市凉州区)。西汉元狩二年(前121年),西汉政府为显示大汉帝国的武功军威,在西北地区原休屠王领地置武
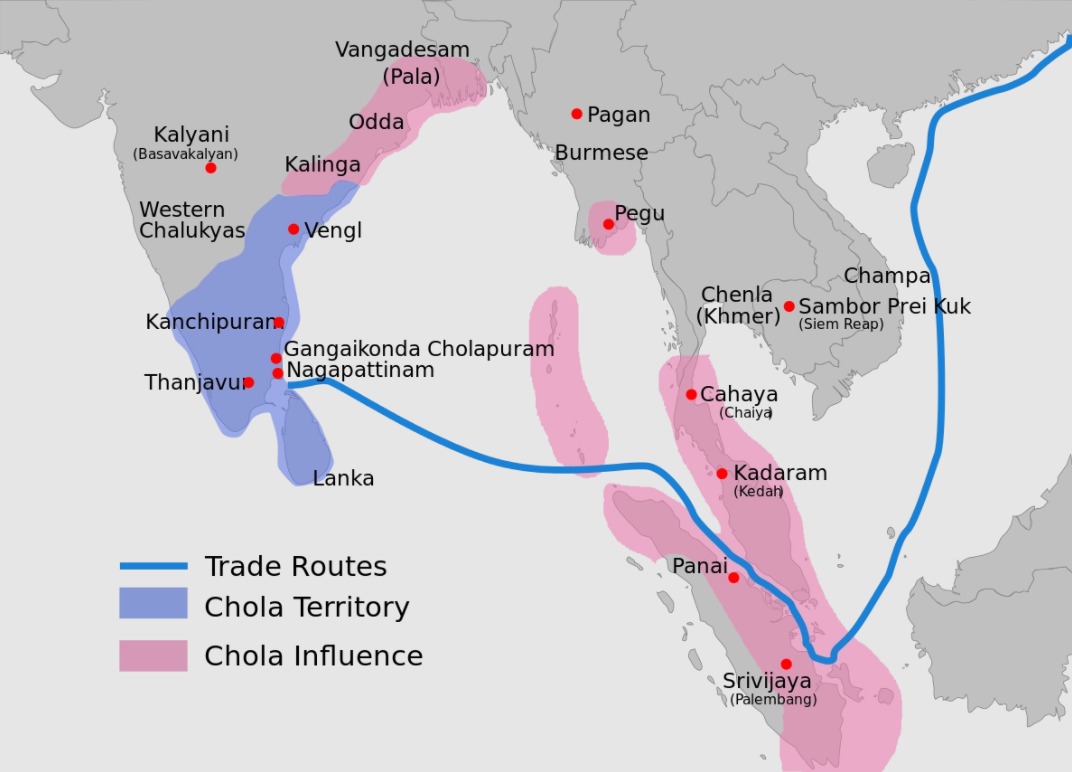
朱罗国(泰米尔语:சோழர் குலம்,宽式IPA:/'ʧoːɻə/),又名注辇,印度半岛古国,其地在今泰米尔纳德邦。建立于公元前三世纪灭亡于公元1279年王朝国祚超过一千五百年。注辇国最早起源于高韦里河(Kaveri)流域,以欧赖宇尔(Urayur)为国都。朱罗王朝的统治者们曾经征服印度半岛南部,吞并斯里兰卡,并占领了马尔代夫,甚至还成功地入侵马来群岛的王国。在公元1010年至1200年期间,注辇国领土南到马尔代夫群岛,最北到安得拉邦的达戈达瓦里河沿岸。朱罗王朝在13世纪进入衰退,随着潘地亚

鲁奇乌斯·多米提乌斯·奥勒里安努斯(Lucius Domitius Aurelianus,通常译作奥勒良,214年9月9日-275年9月),罗马皇帝,公元270年-275年在位。在他的统治期间,收复了罗马帝国曾经失去的三分之二的疆域,将分裂50年的帝国再次统合,使罗马帝国在3世纪末至4世纪初恢复重新统一。他的统治初步解决了罗马帝国的三世纪危机,是几位成功的军人出身的皇帝中的第二位。可惜最后遭到暗杀。

尚可名片
这家伙太懒了,什么都没写!
作者


