蛋白质复性指蛋白质或核酸的天然构象,在分子遭到变性以后,全部或部分恢复。它通常是特指分子生物学中一类生物大分子物质,尤指脱氧核糖核酸(DNA)和核糖核酸(RNA)。指变性的DNA(单链)在适当的条件下,两条彼此分开的多核苷酸链又可以重新结合成双螺旋结构,这一过程称为复性。DNA分子受热变性,双链分开,若再缓慢冷却至室温,则在260nm处吸光度降到变性前的值,这一过程称为“退火”。经退火处理,可使变性DNA的两条多核苷酸链重新聚合成双链螺旋结构,恢复其本来的理化性质和生物学功能。 在变性条件不剧烈,变性蛋白质内部结构变化不大时,除去变性因素,在适当条件下变性蛋白质可恢复其天然构象和生物活性,这种现象称为蛋白质的复性(renaturation)。 蛋白质因受某些物理或化学因素的影响,分子的空间构象被破坏,从而导致其理化性质发生改变并失去原有的生物学活性的现象称为蛋白质的变性作用(denaturation)。变性作用并不引起蛋白质一级结构的破坏,而是二级结构及以上的高级结构的破坏,变性后的蛋白质称为变性蛋白。
精选百科
本文由作者推荐

蛋白质复性
变性蛋白质恢复天然构象等的现象
中文名
蛋白质复性
结果
分子的空间构象被破坏
称谓
变性后的蛋白质称为变性蛋白
影响
因受某些物理或化学因素的影响
蛋白复性
包涵体复性,蛋白质折叠
如果变性条件剧烈持久,蛋白质的变性是不可逆的。如果变性条件不剧烈,这种变性作用是可逆的,说明蛋白质分子内部结构的变化不大。例如胃蛋白酶加热至80~90℃时,失去溶解性,也无消化蛋白质的能力,如将温度再降低到37℃,则又可恢复溶解性和消化蛋白质的能力。
分离步骤
分离包涵体并复性蛋白质的操作步骤并不复杂,从破碎细胞开始,然后将细胞匀浆离心,回收包涵体后,加入变性剂溶解包涵体,使之成为可溶性伸展态,再通过透析等除去变性剂使表达产物折叠恢复天然构象及活性。
但在实际研究中发现,在体外折叠时,蛋白质分子间由于存在大量错误折叠和聚合,复性效率往往很低。究其原因,蛋白质的立体结构虽然由其氨基酸顺序决定,然而伸展肽链折叠为天然活性结构的过程还受到周围环境的影响。
折叠机制
为了有的放矢地开发辅助蛋白质复性的技术,研究工作者纷纷开展了对蛋白质折叠机制的探讨。有两种不同的假设:一种假设认为,肽链中的局部肽段先形成一些构象单元,如α螺旋、β折叠、β转角等二级结构,然后再由二级结构单元的组合、排列,形成蛋白质三级结构;另一种假设认为,首先是由肽链内部的疏水相互作用导致一个塌陷过程,然后经逐步调整,形成不同层次的结构。尽管是不同的假设,但很多学者都认为有一个所谓‘熔球态’的中间状态。在熔球态中,蛋白质的二级结构已基本形成,其整体空间结构也初具规模。此后,分子立体结构再做一些局部调整,最终形成正确的立体结构。总之,蛋白质折叠的具体步骤可用下式描述:
U→I→N
↓
A
即伸展态U经过早期变化成为中间体I,然后由中间体过渡到最后的天然态N。但是从中间体折叠为天然态的同时,另有一条旁路,即中间体相互聚集为凝聚物A(包涵体)。在折叠反应中,从伸展态到中间体的形成是非常快速的,一般在毫秒范围内完成,但从中间体转变为天然态的过程比较缓慢,是反应的限速步骤。当溶液中离子强度或变性剂浓度很低,又无其它辅助手段存在时,聚集趋势占主导地位,导致蛋白质的自发复性效率极低。
一般认为,蛋白质在复性过程中,涉及两种疏水相互作用,一是分子内的疏水相互作用,二是部分折叠的肽链分子间的疏水相互作用。前者促使蛋白质正确折叠,后者导致蛋白质聚集而无活性,两者互相竞争,影响蛋白质复性收率。因此,在复性过程中,抑制肽链间的疏水相互作用以防止聚集,是提高复性收率的关键。
初期成果
80年代后期,人们开展了广泛的蛋白质复性研究。例如,Carlson等发现,单克隆抗体具有协助蛋白质复性的作用;Cleland等发现,向稀释的变性蛋白质溶液中加入适当浓度的聚乙二醇,可抑制蛋白质复性过程中的凝聚沉淀,使蛋白质复性收率提高两倍;Hagen等利用反胶团萃取人工变性的蛋白质后去除变性剂进行复性,复性率可达100%(但由于变性剂的存在,萃取率很低);Batas等利用凝胶过滤色谱介质的分子筛作用可防止变性蛋白质间的接触和凝聚,辅助其正确折叠;Zardeneta等利用去污剂及混合胶团,成功地辅助了脲变性硫氰酸酶复性。
复性研究
环糊精与直链糊精辅助蛋白质复性的研究
1995年,Karuppiah 和Sharma发表文章,介绍了使用环糊精辅助碳酸酐酶B的复性。环糊精由淀粉通过环糊精葡萄糖基转移酶降解制得,是由D-吡喃葡萄糖单元以α-1,4-糖苷键相互结合成互为椅式构象的环状低聚糖,其分子通常含有6~12个吡喃葡萄糖单元。有实用意义的是含6、7、8个吡喃葡萄糖单元的α、β、γ-环糊精,但α-环糊精空腔较小,γ-环糊精价格昂贵,常用的是β-环糊精。环糊精的特征是能形成包络化合物,客体分子从宽口端进入其分子空腔。包络物的形成主要靠非共价键相互作用如范德华力、氢键、疏水相互作用、几何形状匹配等。利用环糊精的疏水性空腔结合变性蛋白质多肽链的疏水性位点,可以抑制其相互聚集失活,从而促进肽链正确折叠为活性蛋白质。
1999年,Sundari等报道了用直链糊精辅助胰岛素、碳酸酐酶和鸡蛋白溶菌酶复性,发现直链糊精基本上能够模拟环糊精在辅助蛋白质复性方面的作用,而且具有这样一些优点:直链糊精的螺旋结构形成一个疏水性空管,可以结合更多的蛋白质分子;在水中溶解度较高,有利于提高复性酶浓度和实验操作;价格比环糊精便宜,实际应用前景更广阔。
分子伴侣
性质近来,越来越多的有关蛋白质折叠的研究已转向利用分子伴侣GroE家族。有些学者已成功地利用分子伴侣在体内和体外辅助蛋白质复性[12、13]。但分子伴侣在实际应用中尚存在费用高并需要与复性蛋白质分离等缺点,因此研究开发分子伴侣的重复利用性及稳定性是实现其应用的关键。
GroEL具有结合蛋白质的作用,相当于变性蛋白质的亲和配基。固定化GroEL柱(固定床)相当于变性蛋白质的亲和吸附层析柱,从而可提高样品的处理量,并使蛋白质在复性的同时得到浓缩和纯化。其特点在于:①不仅可以解决分子伴侣的重复利用问题,而且由于固定床的利用(近于平推流),可提高分子伴侣的利用效率;②由于采用连续操作,原料处理量大,产品可以得到浓缩、纯化;③易于建立相关的模型,对于放大生产具有重要的指导作用。Teshima等利用固定化分子伴侣,在体外辅助了淀粉酶、碳酸酐酶、DNA酶的折叠复性。
还有报道利用“小分子伴侣”对目标蛋白质进行复性[15、16]。“小分子伴侣”是指GroEL的一个片段,而这个片段并不是GroEL的水解产物,而是由基因重组后包括GroEL191-345氨基酸残基片段(16.7kD)或191-376氨基酸残基片段(21kD)密码的基因片段在大肠杆菌中的表达产物。它们能有效地促进亲环蛋白A、硫氰酸酶以及芽孢杆菌RNA酶的复性。由于“小分子伴侣”分子较小,更适合于固定化,且不需另加复性辅助因子(如GroES和ATP等),因此具有很大的应用前景。
人工分子伴侣受蛋白质分子伴侣辅助蛋白质复性的启发,Rozema和Gellman对人工分子伴侣体系(去污剂+环糊精)辅助碳酸酐酶和鸡蛋白溶菌酶复性进行了研究[17、18]。与分子伴侣GroEL+ATP辅助复性的作用机制相似,其复性过程分为两步进行:第一步捕获阶段,在变性蛋白质溶液中加入去污剂,去污剂分子通过疏水相互作用与蛋白质的疏水位点结合形成复合体,抑制肽链间的相互聚集;第二步剥离阶段,加入环糊精,由于环糊精分子对去污剂分子有竞争性吸附作用,去污剂分子被剥离下来,从而使多肽链在此过程中正确折叠为活性蛋白质[17、18]。其反应机制可用下式表示:
U→U-dn→→→U-dn-m→→→F→N
即伸展态U首先与去污剂d形成复合物U-dn,加入环糊精后,小分子去污剂被逐次剥离下来,变为部分折叠、不含去污剂分子的非活性状态F,进而折叠为天然活性蛋白质N。
与GroE等蛋白质分子伴侣相比,联合使用去污剂和环糊精作为人工分子伴侣辅助蛋白质复性具有明显的优点:①人工分子伴侣不属于蛋白质,不易受环境影响而失活,操作条件较为宽松;②去污剂和环糊精均可直接购买,省去分离纯化的步骤;③去污剂与环糊精的分子量较小,容易与蛋白质分离,有利于提高工业生产效率。?
冷冻结晶脱盐冷冻结晶脱盐在蛋白质复性研究方面的一些提示(集中精力在和缓的降低变性盐浓度上,之前的所有方法基本都是用稀释液体去增大体积,这个方法是让变性盐脱离溶液……)。
通常意义上人们普遍认为冷冻是一种变性因素,原因是在溶液的冻结过程中会伴随发生物态的巨大变化,进而导致蛋白质分子表面的离子氛围、pH、水化状态等各种因素的改变,因此变性也就是自然而然的结果。不过从事生物领域的多半都有冻存细胞的经历,由此可以得出结论,在合适的防冻剂存在的条件下,蛋白是可以耐受低温的。或者至少能有相当的蛋白能在低温下保持活性。况且在变性液体中也不存在有活性的蛋白质,本身都是以
自由肽链
的形式存在,因此冷冻对肽链的损伤似乎可以不用考虑(对此还需要进一步的研究)。换个角度从物理学方面看,“冰”是一种 单矿岩,其生成过程中会自动排除其中的杂质。这也就是为何海上的冰山都是淡水的缘故。通常人们用于溶解包涵体肽链的液体是高浓度的变性盐(一般是8M的尿素,或者6M的盐酸胍,两种盐的密度都在1.3左右),而这种溶质的溶解度受温度影响很大。随着温度的降低,会发生饱和、过饱和、结晶析出现象。溶质结晶析出自然也就意味着溶液浓度的下降,进而使变性能力的下降。下降到一定程度时,肽链其内在的塌缩的趋势逐渐占据优势,引导肽链进行自行复性至其活性状态,即能级最低状态。也就是说,肽链折叠的
动力
是由肽链的一级结构(氨基酸序列)决定的,人类直到所能做的复性过程其实质不过就是提供了一个变性能力和缓下降的变性体系
(由尿素或者盐酸胍的浓度下降来实现)。在这个过程中最需要防止的就是肽链之间的错误折叠(链间形成次级键),这会产生“雪崩样”的效果,导致大量多聚体沉淀(密度也是1.3左右)。冷冻优势
冷冻方式用于蛋白复性可能的优势有几个方面。一、这种方式的简单易行,只需要冷冻设备即可;二、此方法的运用可以导致溶液体积的减少,很大程度上对蛋白进行浓缩,方便下一步的处理;三、变性盐以结晶沉淀方式脱离溶液,防止大量高污染复性尾液的产生;四、此方法的体系完全开放,可以很好的对接其他复性方式。至少冷冻一下降低一些初始变性盐浓度也是相当有价值的;五、此方法即便失败也几乎没有任何损失,体系可以很方便的复原,这一点是其他任何方法都不具备的,极其适合科研探索。

蛋白质复性

蛋白质复性

蛋白质复性

蛋白质复性
6、蛋白质的高压冷结晶复性法:
如果把蛋白复性仅只理解为肽链在不同变性盐溶液浓度中的存在状态。那么蛋白复性就没什么难的,无非是把尿素浓度从4M平缓的降低到2M即可。冷结晶析出无疑找到了一条行之有效的途径。
但是之前的冷结晶方法似乎有个硬伤,也就是2M尿素时的溶解度,有可能相对的温度在零下——液体冻结对蛋白活性的实质伤害。解决办法可以加入甘油之类的防冻剂降低水的冰点。另外如果将结晶物理学的一些原理加于其中应该即可解决这个问题。

蛋白质复性
在物态的固液汽三相变化中,水是种异类。其在零上4度的液体时拥有最大密度。固体变为冰时,其体积反倒变大,密度变小。在高压强下,水的冰点下降。冰雪踩上去打滑就是冰在高压强下融化的日常例子。而尿素之类的变性盐其固体密度大,高压强更能促进它们结晶析出。只是还不知道能以何种的压强达到目的,希望不会有太高的要求。更有可能的是辅助以防冻剂来达到所需效果。
蛋白质复性相关的文章

蚜虫(英文名:Aphid),是球蚜总科(Adelgoidea)和蚜总科(Aphidoidea)昆虫的统称,又称腻虫、密虫,隶属于节肢动物门昆虫纲半翅目,包括球蚜科、根瘤蚜科、纩蚜科、平翅绵蚜科、扁蚜科、瘿绵蚜科、群蚜科、毛管蚜科、斑蚜科、毛蚜科、大蚜科、短痣蚜科和蚜科。蚜虫体长0.5~7.5mm,大
疮疡是指各种致病因素侵袭人体后引起的一切体表化脓感染性疾病的总称,包括急性和慢性两大类。中国古代用以泛指多种外科疾患,后世将外科分为疮疡与杂证两大类。疮疡多由毒邪内侵,邪热灼血,以致气血凝滞而成。其包括了体表上的肿疡及溃疡、痈、疽、疔疮、疖肿、流注、流痰、瘰疬及有关皮肤病的内容,是中医外科疾病中最常

双子叶植物(学名Dicotyledons)[1],旧称双子叶植物纲、木兰纲。属种子植物门,是指一般其种子有两个子叶之开花植物的总称,约有199350个物种。现在双子叶植物不再被视为是一个恰当的类群名称,且此一名称亦至少不被使用在分类的意涵上。但前述之双子叶植物的绝大部份可以分至一名为真双子叶植物分支的单系群。此一单系统可以以其花粉的结构于其他的开花植物所区隔。其他的双子叶植物和单子叶植物的花粉都是单沟或单沟衍生的样式;而真双子叶植物分支的花粉则为三沟或三沟衍生的样式,其花粉的沟上会有三个或三个以上的细孔。
路易斯,美国物理化学家。从他的电子理论出发,他还为酸碱作出了更为广泛的定义:可接受一个电子对的物质是酸,可给出一个电子对的物质是碱,形成路易斯酸碱理论。常见的路易斯酸有氯化铝、氯化铁、三氟化硼、五氯化铌以及镧系元素的三氟甲磺酸盐。路易斯酸,又称亲电子试剂,指可以接受电子对的物质,这是根据路易斯的酸碱电子理论对酸的定义确定的。由于它所包含的物质极为广泛,也称广义酸。
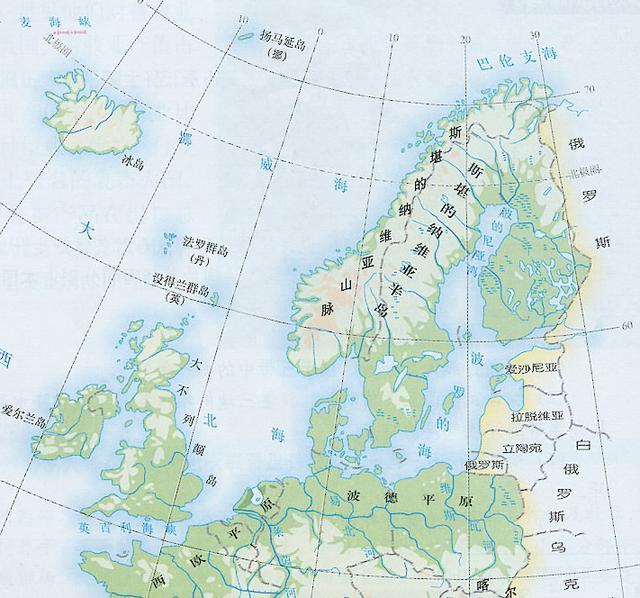
斯堪的纳维亚半岛(丹麦语:Skandinaviske halvø,瑞典语:Skandinaviska halvön,挪威语:Den skandinaviske halvøy,芬兰语:Skandinavian niemimaa),位于欧洲西北角,其濒临波罗的海、挪威海及北欧巴伦支海,与俄罗斯和芬兰北部接壤,北至芬兰,意为“斯堪的纳维亚人居住之地”。“斯堪的纳维亚”(Scandinavian)一词源自条顿语“skadino”,意为“黑暗”,再加上表示领土的后缀-via,全名意为“黑暗的地方”。因半岛 地处高纬

尚可名片
这家伙太懒了,什么都没写!
作者