
可可肿枝病毒是学名 Cacao swollen shoot virus,异名是可可病毒1号。
精选百科
本文由作者推荐

可可肿枝病毒
可可肿枝病毒
中文名
可可肿枝病毒
正文
可可肿枝病毒
是学名 Cacao swollen shoot virus,异名是可可病毒1号 Theobrama virus 1 (Rev. appl. Mycol. 27: 178);Marmor theobromae (Rev. appl. Mycol. 27: 178);Theobromavirus inflans (Rev. appl. Mycol. 27: 178);Cacao swollen shoot virus Marmor theobromae;可可病毒1号 Cacao virus Ⅰ;Cacao swollen shoot badnavirus;Cocoa swollen shoot badnavirus,英文名 Cacao swollen shoot virus。分类地位
花椰菜花叶病毒科Badnavirus 属可可肿枝病毒。
分布
CSSV在非洲西部广泛分布如加纳、象牙海岸、尼日利亚、多哥、塞拉利昂、斯里兰卡。CSSV在亚洲锡兰、印度和马拉西亚、沙巴也有发生。西半球:哥伦比亚、特立尼达、多巴哥和委内瑞拉。
寄主植物
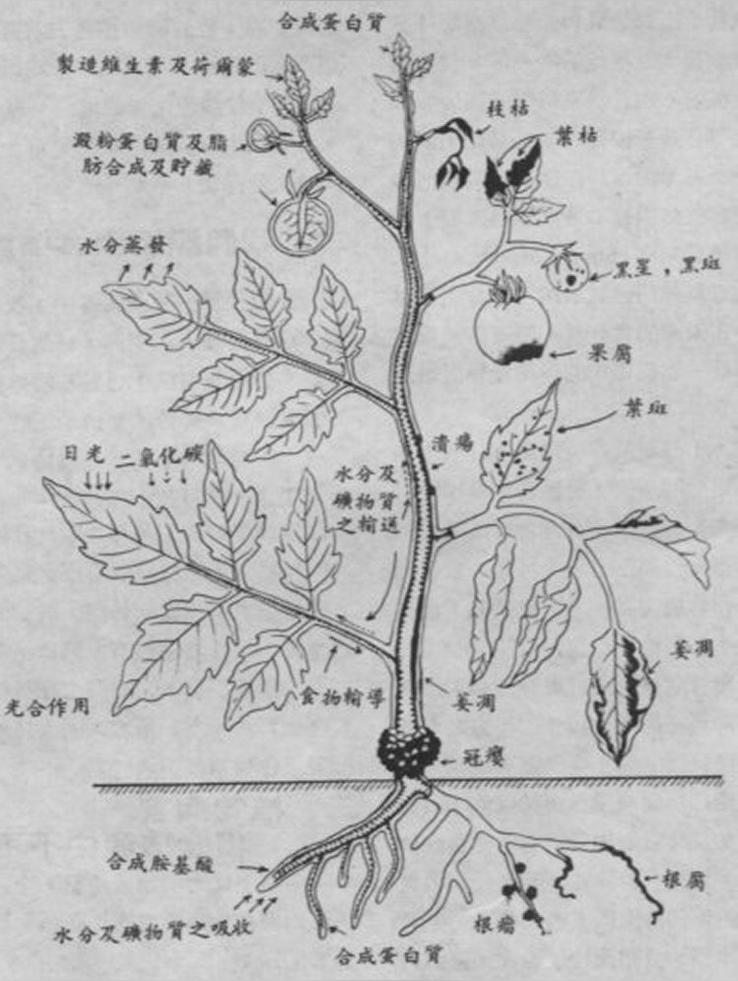
(图)可可肿枝病毒
试验寄主
只局限于木棉科、椴树、梧桐科、锦葵属中30种植物(Posnett, Robertsin, Todd,1950; Tinsley & Wharton, 1958)。自然寄主仅限于可可、Cola chlamydantha, Ceiba pentandra, Cola gigantean var. glabrescens(Posnette,1950)和苹婆(Sterculia tragacantha)(Legg & Agbodjan,1969)。
诊断寄主
可可(Theobroma cacao)上的症状:茎肿大,幼叶沿叶脉变红,老叶脉间褪绿。
黄麻(Corchorus spp)上的症状:黄麻是高感寄主。许多株系感染后,黄麻死亡(Tinsley & Wharton, 1958)。一般株系侵染后,寄主不表现症状。病毒感染吉贝(Ceiba pentandra)、Cola chlamydantha和Cola gigantea后,引起暂时叶脉间褪绿;但感染猴面包树(Adansonia digitata)引起永久褪绿和严重矮化(Posnette et al., 1950)。
繁殖寄主
可可、猴面包树、木棉。
测定寄主
可可树(计算幼苗发病率)。
危害情况
CSSV在西非引起重大经济损失,有许多变种。寄主范围窄,由14种粉蚧传播,也能机械传播到可可豆。CSSV为杆状颗粒,大小为121-130×28纳米。
许多株系在可可上产生症状明显,并使可可减产,但不能致死。除非先用弱毒株系侵染可可,再用强毒株系如Calonectria rigidiuscula攻毒(Thresh, 1960)。一些无毒株系只在小范围内爆发,通常只引起茎肿大,对植物生长及产量几乎没影响。有些病毒株系是广泛分布且是致死株系。在非洲西部the severe New Juaben株系引起Amelonado可可茎和根部明显肿胀(Fig. 3、Fig. 4)、测根坏死(Fig.4)、落叶,产生小环斑斑驳豆荚(Fig.6)。结的豆荚又少又小,新叶产生短暂的叶斑变红现象,老叶产生褪绿斑和蕨叶等(Posnette, 1947)(Fig.1, Fig.2)。CSSV侵染一年后,减产25%,二年后减产50%,3-4年后植株死亡(Crowdy & Posnette, 1947)。
形态特征
粒子特性
沉降系数(S20, w):218S。
粒子结构
粒子杆菌状(Fig. 5),大小121-130×28纳米。在中性2%钾磷钨酸溶液中,无明显亚结构(brunt, 1964; Kenten & Legg, unpublished)。
粒子组分
未知。
生物学
血清学
效价为1/128的抗血清用来检测来自New Juaben, Mampong和Anibil病毒株系。低效价可能是由于免疫时相对低的病毒含量造成的凝胶扩散试验用于检测病毒。
关系
New Juaben, Mampong和Anibil三个株系间存在抗原差异,但三者之间及与可可斑驳病毒间有一定联系(Kenten & Legg, unpublished)。
病汁液稳定性
粗汁液不具侵染性,但病毒抽提液加抗氧化剂后侵染性增加5%(Brunt & Kenten, 1962)。用病毒浓缩液,传毒率能增加60-90%。致死温度:55-60℃(10分钟),稀释限点10-3-10-4,体外存活期为2-3个月(2℃)(Brunt & Kenten & Nixon, 1964; Kenten & Legg, 1965)。
病毒纯化
因病叶含粘液和多酚,抽提纯化较难。提纯方法:病叶加10-20倍体积的0.05M磷酸缓冲液(PH 6.0-8.0)加蛋白(1-2% w/v 白蛋白或牛血清白蛋白或皮粉)再加0.05M巯基乙酸或者0.01-0.005M二乙基二硫氨基甲酸酯匀浆(Brunt & Kenten, 1963),不同速度分次离心沉淀病毒或者先硫酸铵沉淀再超速离心(75000 g/hr)沉淀病毒。
病毒与细胞、组织的关系
病叶褪绿部分的组织不分化,细胞间隙小,内含小而扁平已退化叶绿体。在茎和根部肿大区次生木质部和韧皮部增加,但没有韧皮部坏死现象发生(Knight & Tinsley, 1958)。
传播途径
介体传播
至少由14种粉蚧可传播CSSV。虽然在尼日利亚,Planoceccoides njalensis, P. Kenyae和P. citri同样多,但P. njalensis是主要的传播介体。1龄、2龄、3龄若虫和成雌虫能传毒,成雄虫不能传毒。粉蚧饲毒前饥饿时间的延长有助于传播效率的增加。最短获毒饲育期为20分钟,长至50分钟(Posnette & Robertson,1950),最长可达2-4天(Roivainen,1969)。没有明显的潜育期,15分钟后昆虫就能传毒,最长为2-10小时(Posnette & Robertsin, 1950; Dale, 1955)。病毒最昆虫体内的持久性为3小时或少于3小时(Posnette & Strickland, 1948),但饥饿成虫和1龄若虫分别能持毒49和24小时(Lister, 1953)。卵不能传毒。
介体特异性
Ferrisia virgata能传播所有株系,除了可可斑驳病毒由Pseudococcus longispinus传毒(Posnette,1950)。
种传
The New Juaben和Bisa株系不能种传(Posnette 1947)。
检疫与防治
检疫措施
1. 禁止从疫区引进可可;
2. 特殊需要的,经指定机构审批同意,可引进少量可可并附出口国检疫证书;
3. 限制引进数量,建议每个品种5—10个可可;
4. 引进可可必须在隔离检疫苗圃或隔离温室中隔离试种1—2年,确为无毒,方可放行;
5. 引进的优良品种资源,种质可利用茎尖脱毒获得无毒苗保存和传递。
检疫方法
1. 种植观察:观察植株茎干部症状;
2. 鉴别寄主反应
3.血清学反应:琼脂免疫扩散测定
4.细胞组织病理学检验
可可肿枝病毒和可可斑驳病毒都由粉蚧传播(Posnette 1947),病毒粒子和特性相似(Brunt,1964; Kenten & Legg,1967),血清学存在差异(Kenten & Legg, personal communication)。目前学术界未确定可可斑驳病毒是一种新病毒还是可可肿枝病毒的一种血清型。
在特力尼亚,由粉蚧传播的两个不同病毒株系侵染可可;在夏威夷,粉蚧传播的另一种病毒侵染凤梨(Carter,1963);对这些病毒知之甚少。又有报道称:病毒侵染印度尼西亚、哥伦比亚、多米尼亚共和国和委内瑞拉的可可,这些都为可可肿枝病毒和可可斑驳病毒之间的比较带来困难(Thresh & Tinsley, 1959)。
病害防治
1. 检疫
严禁从发病国家引进可能携带CSSV的种子、苗木和繁殖材料。在进口材料中一旦发现CSSV,植株全部销毁,以防蔓延,特殊需要的需办理特许审批手续。
2. 消除生产区传毒介体——粉蚧。
3. 组培脱毒生产无毒苗,种植无毒苗。
可可肿枝病毒相关的文章

小编整理:马涛,湖北省委原常委、省军区原司令员。马涛,男,汉族,1961年6月生,中国共产党党员,中国人民解放军少将军衔。曾任湖北省委常委、省军区司令员。湖北省国防动员委员会副主任。马涛湖北省军区原司令员马涛,男,1961年6月生,中共党员,中国人民解放军少将
小编整理:平冈佑太是一位日本男演员,出生于1984年9月1日,来自日本山口县。他在2002年参加JunonSuperBoyContest并获得冠军,从而开始进入娱乐界。以后来,他出演了多部电视剧,包括《借着雨点说爱你》和《东京friends》等。而在电影方面,
萨拉丁·本(1137年—1193年),全名阿马里克·纳赛尔·萨拉丁·阿布一穆沙发·优素福·伊本·阿尤布(Salah-al-Din Yusuflbn-A yyub),库尔德人,埃及阿尤布王朝的开国君主,在位23年。

尚可名片
这家伙太懒了,什么都没写!
作者

