
精选百科
本文由作者推荐
棕色脂肪组织
哺乳动物体内呈棕色的脂肪组织

生物解释
人和哺乳动物体内的脂肪组织可分为白色脂肪组织和棕色脂肪组织。前者就是我们通常认识的那种脂肪,广泛分布于皮下组织和内脏周围。棕色脂肪组织存在于人和所有哺乳动物体内,但主要是在新生儿和幼小的哺乳动物体内。
这两种脂肪组织与白色脂肪组织既然都叫“脂肪组织”,它们的共同点当然就是细胞内都含有脂肪。它们的区别在于,棕色脂肪组织的细胞体积较小,细胞中脂肪颗粒较小,却含有大量线粒体,细胞周围有丰富的毛细血管,交感神经纤维直接到达棕色脂肪组织的细胞膜上。
外表不同、结构不同,它们在能量代谢中所起的作用也截然相反。白色脂肪组织是能量仓库,它的作用是将多余的能量以脂肪的形式储藏起来;棕色脂肪组织则是通过它细胞内大量的线粒体将食物中的能量转化成热能。棕色脂肪细胞的线粒体有一种称为解耦联蛋白1(UCP1)的物质,这种物质使得葡萄糖和脂肪酸分解产生的能量不能转化为ATP(为生物体直接供能的物质),而只能转化为热能。这就是为什么棕色脂肪组织较多存在于新生儿和幼小哺乳动物体内的原因:因为小家伙们刚生出来,身体的调节功能还比较差,不能通过“打哆嗦”(肌肉颤栗)这种方式产热来抵御外界的寒冷,所以需要棕色脂肪组织的帮助。打个比方来说,白色脂肪组织就好比被子,可以起保暖的作用;棕色脂肪组织就好比电热毯,通过产热来帮助机体抵御寒冷。
功能
分布概况
1BAT的概况
人和哺乳动物体中的脂肪组织分为棕色脂肪组织(BAT)和白色脂肪组织(WAT)。BAT因其细胞内含有大量血红蛋白和高水平的血红素卟啉,细胞呈棕色而得名。BAT在所有哺乳动物体中都存在,主要是幼小哺乳动物体中,而在成熟的哺乳动物体中仅有少量存在。BAT细胞体积比WAT小,细胞表面密布交感神经纤维,细胞中脂肪颗粒很少,此种组织细胞内含有大量线粒体,细胞周围含有丰富毛细血管,这与其产热功能相匹配。WAT与BAT在能量代谢中发挥着截然相反的作用。WAT主要是通过甘油三酯贮存能量,而BAT则通过产热来消耗能量。早在1961~1964年,人们就发现啮齿动物能利用体内棕色脂肪组织,对寒冷刺激、休眠、采食诱导,营养不足(低蛋白质)时,产生热量维持体温十分重要(Smith等,1961)。有研究发现BAT在成年人或成年动物体中已经消失,但近年来的研究表明,户外工人和长期饮酒人群的颈动脉周围和心包存在BAT,成人的一些部位的脂肪组织检测到了UCP1(解偶联蛋白1)及其RNA,还有一些研究指出,成人脂肪组织中有大量的棕色脂肪细胞分布并表达UCP。
非颤栗性产热(NST)对哺乳动物在冷环境下的生存是必需的,棕色脂肪组织是哺乳动物体内非颤栗产热的主要来源(Smith等,1969;Nicholls等,1984;Himms等,1990)。对于哺乳动物和人,棕色脂肪组织主要分布在:①肩胛间区;②腹部大血管及周围;③肌肉、颈部血管及肾周、胸部动脉和下腔静脉周围分布小块棕色脂肪组织。这种组织的主要功能是通过非颤栗产热维持动物的体温及能量平衡,BAT对于维持动物的体温和能量平衡起重要作用,对幼龄哺乳动物尤为重要(Himms等,1990;Jansky等,1995)。
产热机制
2BAT产热机制——去甲肾上腺素(NE)控制产热
哺乳类动物BAT活动的最终目的是产生热量。BAT主要作用是调节机体温度,参与能量的消耗,因而与保持机体重量也有关。
2.1去甲肾上腺素对BAT的快速作用——控制产热
去甲肾上腺素是交感神经的主要递质,冷暴露条件下,交感神经末梢释放NE激活组织,是产热增加的生理基础(HimmsHagen,1990;Baumuratov等,2003;Baumuratov等,2005)。寒冷刺激,脂肪细胞周围交感神经末梢释放的去甲肾上腺素,通过作用于棕色脂肪细胞上的受体,使细胞内cAMP浓度发生改变,最终影响甘油三酯分解为甘油和脂肪酸。
机体中的肾上腺素能受体(AR)有α、β两种,其中β分为β1、β2、β3,β1在成熟的棕色脂肪组织中表达;β2在棕色脂肪组织中没有表达;β3在棕色脂肪组织和白色脂肪组织中都有表达。每种受体的作用方式和机理不同。
NE信号主要通过β3肾上腺素能受体控制产热(HimmsHagen等,1990;Smith等,2004;Oana等,2006),NE通过作用于棕色脂肪细胞上的β3肾上腺素能受体,与Gs蛋白偶联活化腺苷酸环化酶(AC),使细胞内cAMP浓度增加,从而激活cAMP依赖的PKA,后者使敏感脂酶磷酸化,最终使促进脂滴中贮存的甘油三酯分解为甘油和脂肪酸(见图1);PKA使cAMP反应元件结合蛋白(CREB)磷酸化,成为有活性的形式,有活性的CREB能直接诱导PPAR激活UCP1和PGC-1α的表达。这种生热主要是通过脂解作用激活了UCP1,是大多数哺乳动物的主要生热方式。但是对于像猪这样的BAT中缺乏β3肾上腺素能受体的动物的产热方式则主要是通过β1肾上腺素能受体的作用(Atgie等,1996;Himms-Hagen等,1995)。NE刺激BAT,β1肾上腺素能受体表达增加,使交感神经紧张,从而产热。α2肾上腺素能受体抑制产热(McMahon等,1982),NE通过作用于棕色脂肪细胞上的α2肾上腺素能受体,与Gi蛋白偶联,抑制腺苷酸环化酶(AC),使细胞内cAMP浓度降低,抑制产热(见图1)。α1肾上腺素能受体作用方式:NE刺激或冷暴露,α1肾上腺素能受体增加,使线粒体内Ca2+浓度增加,增加产热,这种产热可能还是与cAMP有关(Zhao,1997;Baumuratov等,2005)。Ca2+在此扮演着第三信使的作用,类似于cAMP(Baumuratov等,2005)。
2.2去甲肾上腺素对BAT的慢性作用
去甲肾上腺素对BAT的慢性作用主要是引起棕色脂肪细胞的增殖和分化。
2.2.1细胞增殖
1964年Camerond等就发现动物在寒冷环境中BAT分裂指数增加,10d左右细胞内DNA含量可增加3~4倍。给小鼠和大鼠注射NA(核酸)也可产生类似的变化(Rehnmark等,1989;Geloen等,1990)。为了进一步研究这个过程,Bronnikov等(1992)用原代培养的棕色脂肪细胞进行研究。细胞在含有小牛血清的培养基中可自然分裂,通常在6d左右就可达到融合。在细胞融合前的增殖阶段,NA可明显增加DNA合成。在无小牛血清环境下,培养细胞不出现自然分裂,此时NA也能明显刺激DNA合成。这表明NA确实具有促进棕色脂肪细胞增殖的作用。一些能促进细胞内cAMP浓度增高的物质都可通过与NA类似的方式增加DNA合成。因此可以说cAMP是非成熟细胞增殖的信使。
2.2.2细胞分化
在生理性诱导的募集反应中,也出现细胞分化程度的增高,表现在线粒体生成增多和UCP基因表达增加,这些变化可增加组织的产热能力。细胞分化的调节也可用培养细胞来研究。NA可增加UCPmRNA的表达和细胞内UCP的含量,这种刺激反应在细胞融合期最为明显(Rehnmark等,1990)。利用能增高细胞内cAMP浓度的物质(如forskolin或cAMP的类似物)也能引起相同的反应,很明显NA引起基因表达主要是由β-AR介导。因此在成熟棕色脂肪细胞,NA通过激活β3-AR和cAMP增高诱导基因的表达,cAMP也是棕色脂肪细胞分化的信使。cAMP引起非成熟细胞DNA的合成,在成熟细胞中则引起UCP的表达。
解偶联蛋白
3BAT与解偶联蛋白
线粒体内膜的解偶联蛋白(UCP)是决定BAT功能的关键因素(Heaton等,1978)。UCP是参与能量代谢的重要蛋白质。解偶联蛋白是一种在棕色脂肪组织特异表达的线粒体内膜蛋白质,分子量为32kDa。UCP的作用是作为脂肪酸阴离子转运载体。
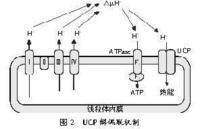
电子传递链产生H+跨线粒体内膜的势能。在偶联状态下,势能驱使H+通过ATP合成酶而重新回到线粒体基质中,同时将势能转化为ATP的化学能。由于UCP在线粒体内膜上有特殊的通道,由电子传递链泵出的氢离子直接通过这种通道流回线粒体内,使跨线粒体内膜的质子电化学梯度消失,ADP不能进行磷酸化形成ATP,物质氧化与ATP生成脱偶联,即质子渗漏,而成为产热过程,能量以热能形式释放,减少了ATP合成(Lowell等,1997;Boss等,2000)(见图2)。
3.2UCP分类
UCP有五大类,UCP1主要存在于棕色脂肪组织中,UCP2广泛分布于组织器官中(包括棕色和白色脂肪组织),UCP3分布在骨骼肌和BAT中,UCP4和UCP5主要分布在中枢神经系统中。在棕色脂肪组织中存在的只有UCP1、UCP2和UCP3。UCP1在啮齿类动物中的主要作用是产生热量以维持体温及能量稳态,在控制体重和脂肪含量中有重要作用;UCP2主要阻止脂肪酸氧化和脂质的累积(Yu等,2000);UCP3非颤抖的生热作用(Jezek,1998)。近年来的研究表明,UCPs不仅与组织的产热有关,而且关系到个体的代谢方向与速率(Nisoli等,1998)。UCP2、UCP3基因突变小鼠表现出肥胖与糖尿病;敲除UCP1基因的小鼠虽不表现为肥胖,但表现出明显的畏寒倾向(Nisoli等,1998)。
3.3UCP的主要生理作用
3.3.1调控线粒体合成ATP与产热
UCP具有解偶联活性,能降低线粒体膜电位,限制ATP合成,增加产热,调节机体产热,维持体温。UCP介导H+内流,使能量以热量的形式散发,有助于维持机体体温。UCP1主要分布于棕色脂肪组织(BAT)中,BAT是产热的主要组织。通过基因敲除技术,使小鼠UCP1基因失活后,其对寒冷不耐受(Nisoli,1998)。
当能量底物供应过量、ATP水平增高或当ADP不足时,线粒体呼吸受到抑制,NAD+/NADH比例降低,UCP使线粒体膜电位低于某一阈值,限制ATP的产生,促进细胞呼吸,维持NAD+/NADH比例,对合成脂肪、生酮及氨基酸代谢均有重要作用。
3.3.3抑制ROS(活性氧)的生成
ROS的产生与△μH+呈正相关,线粒体外膜的质子回流减少,引起△μH+提高,导致线粒体呼吸链的电子传递阻力增大,抑制电子沿呼吸链传递,使电子易发生泄漏而产生O2-·。这样当ADP下降,ATP合成减少、△μH+提高时,ROS产生增加,导致氧化应激增强(Fleury等,1997)。脂肪酸β氧化增强时,UCP2和UCP3的mRNA水平升高。脂肪过度氧化使ROS过量产生,UCP2、UCP3可能会通过解偶联作用,防止△μH+的过度提高,使ROS产生减少。UCP2和UCP3在脂肪酸代谢中的作用可能也是减少脂肪酸过度代谢产生的ROS。此外寒冷会增加ROS的产生,而寒冷也会诱导UCP1的表达,所以寒冷增加UCP1来产热,可能只是UCP1抑制ROS的副作用(Fleury等,1997)。因此可以认为UCP最直接、最本质的功能可能还是抑制ROS,而其它的功能只是这一功能所产生的结果(Fleury等,1997)。
PGC1α关系
4BAT与PGC-1α
过氧化物酶体增殖物激活受体γ辅助激活因子(PGC-1α)是诱导棕色脂肪细胞UCP1高表达的重要激活因子。PGC-1α主要在棕色脂肪细胞表达,而白色脂肪细胞表达极少(Liu等,2004)。PGC-1α与棕色脂肪细胞的分化及其生理功能关系密切,是调控机体能量代谢的重要基因。在调节肝糖异生、骨骼肌葡萄糖转运、胰岛素敏感性、适应性产热、线粒体功能、脂肪酸β氧化、脂肪细胞分化、胆固醇逆向转运等方面发挥重要作用(Michae等,2001;Boustead等,2003;Louet等,2002)。
4.1PGC-1α与能量代谢的关系
在寒冷、有氧运动和禁食条件下,交感神经兴奋诱导BAT内PGC-1α表达,提高UCP1水平,从而增加了机体能量消耗(Pierre,2003)。敲除棕色脂肪细胞PGC-1α基因后,其线粒体呼吸链功能酶及脂肪酸氧化功能明显降低;冷刺激下,PGC-1α基因敲除鼠UCP1停留在基础水平(Nedergard,2001)。遗传引起的肥胖症,比如ob/ob、db/db、fa/fa肥胖大鼠,其棕色脂肪细胞的PGC-1α及UCP1含量明显减少,脂肪酸β氧化的酶活性降低,能量代谢率下降。可见棕色脂肪细胞PGC-1α及UCP1表达异常参与了肥胖症的产生(Prpicv,2003)。
4.2PGC-1α诱导脂肪代谢和细胞分化
在培养的小鼠初代肝细胞中瞬时转染PGC-1α可激活肉毒碱棕榈酰基转移酶Ⅰ(CPT-I,脂肪酸β氧化的限速酶)基因的表达,激活肝脏脂肪酸β氧化(Louet等,2002)。当PGC-1α被导入白色脂肪细胞后,从而使WAT细胞具有BAT细胞的特征,能诱导UCP1基因的表达和线粒体的合成,后两者决定着棕色脂肪细胞特异性分化(Tiraby等,2003)。PGC-1α是肾上腺素信号途径的下游靶点,而后者是决定体外培养细胞在寒冷刺激下棕色脂肪细胞定向分化的关键调节因子。PGC-1α能诱导WAT向BAT转化,促进BAT产热耗能。因此,定向诱导PGC-1α在WAT表达,增加白色脂肪细胞内脂质氧化,加大机体能量消耗,可能是进一步研究减肥治疗的一个方向。
4.3PGC-1α促进BAT和骨骼肌细胞产热
影响因素
5影响BAT含量及功能的因素
5.1遗传的因素
不同的动物品种BAT在代谢上有差异。Smith等(2004)用NE灌注Brahman(B)(高死亡率品种)和Angus(A)两个品种牛的BAT组织,结果发现高死亡率B品种BAT中β受体的含量是A品种的2~3倍;用NE灌注两个品种牛的BAT组织,用棕榈酸做合成脂肪的原料,发现BAT中脂肪的生成A显著高于B;出生后冷暴露48h,B品种的BAT收缩,而A品种没有,B品种的BAT很快耗尽;体外培养,发现A品种皮下BAT先于B发育,说明A品种有较大的产热能力。可以看出高死亡率B品种BAT分解旺盛,但不持久,脂肪的合成能力差,导致冷适应下死亡率高(Landis,2002)。
随年龄的增加,BAT减少,刚出生的婴儿,棕色脂肪组织约有100g,然而到了青春期左右开始减少,最后减至40g左右,但并不会完全消失。在胚胎发育的不同时期,BAT发育也有差异。特别是在出生前14~24d,BAT细胞的体积增大,分娩前的棕色脂肪细胞,线粒体拉长,且高度盘绕,嵴高度分化,BAT中线粒体增加,UCP的表达增加。BAT的形态学和UCP的表达变化为胎儿脱离母体做准备。
5.2季节和气温
季节和气温影响动物BAT的含量。Dew等(1998)对一种冬眠的小型哺乳动物研究发现BAT细胞中线粒体冬天比夏天多,脂肪滴冬天比夏天小。羔羊冷暴露48h,BAT中脂肪全部耗尽(Landis,2002)。
5.3营养因素
5.3.1能量
高能日粮可使BAT基因表达和沉积发生变化,促进UCPmRNA的表达(Surwit,1998),显著增加大鼠BAT的重量和降低血清甘油及甘油三酯浓度。高能日粮调控BAT-UCP1mRNA、UCP3mRNA的表达机制一样,可能是影响β3肾上腺激素(Savontaus,1998)、瘦素(Scarpace,1998)、甲状腺激素(Larkin,1997)和寒冷刺激(Boss,2000)对UCP1mRNA、UCP3mRNA的调控作用,显著增加BAT-UCP1mRNA、UCP3mRNA水平,但不影响UCP2mRNA的表达。高能日粮增加脂肪沉积,促使血清瘦素升高(Javier等,2001),抑制食物食入和增加能量消耗,促进BAT-UCP1mRNA的表达(Scarpace等,1998)。Marcia等(1992)发现,饲喂高蛋白质和高碳水化合物的日粮,BAT的产热能力下降。
喂养含有较高水平多不饱和脂肪酸的高脂饮食导致棕色脂肪产热活性增加及BAT-UCP1mRNA表达增加,而且富含n-3多聚不饱和脂肪酸(PUFA)日粮比富含n-6PUFA的日粮增加BAT-UCP1mRNA表达(Takahashi,2000),使BAT-UCP基因表达增加2~3倍(Takahashi,1999)和BAT-UCP1、BAT-UCP2分别增加4倍、3倍(Rippe等,2000)。但用熟猪油喂养成年大鼠4周,结果发现大鼠饮食量略有下降,体重无明显增加;肩胛间棕色脂肪体积无明显变化,颜色无改变;对棕色脂肪线粒体上UCP无明显影响。可能的原因是:①多不饱和脂肪酸比饱和脂肪酸能强烈地刺激交感神经系统;②改变饮食中多不饱和脂肪酸水平,可能改变了膜的功能,如受体-酶偶联和激素敏感性,多不饱和脂肪酸对偶联程度有重要影响;③多不饱和脂肪酸可能直接影响棕色脂肪,如亚油酸可能优先运输到棕色脂肪,通过脂肪酸急性影响质子通道,导致线粒体氧化呼吸解偶联增加(Matsuda等,1997)。Encinias等(2004)给怀孕50d的母羊补饲亚油酸,提高羔羊的成活率2.9%(P=0.03),但不提高断奶重和BAT的沉积。
限食或饥饿会降低BAT的活性,可能的原因是降低了交感神经的刺激,降低了UCP的表达(Young等,1982;Rothwell等,1982)。给小鼠或大鼠饮用酒精水可以增加BAT的含量(Huttunen等,1988、1990;Elsukova等,2001)。
5.3.2蛋白质
给小鼠或大鼠饲喂低蛋白日粮可以增加BAT募集,UCP表达增加(Glick等,1981、1983、1984、1985;LeBlanc等,1997;Specter等,1995)。而饲喂高蛋白质和高碳水化合物的日粮,BAT的产热能力下降(Marcia等,1992)。可能是因为低蛋白日粮,增加了动物的采食量,导致动物肥胖,使机体中瘦素含量增加,增加了对棕色脂肪调控中心的刺激,从而增加了BAT募集,使能量的利用效率下降,反过来抑制肥胖的发生。
5.3.3微量元素
适量补锌可以通过增加棕色脂肪组织线粒体中UCP含量,提高机体的耐寒力。补锌以及冷暴露未改变大鼠血浆锌水平,但可以使肝脏和肌肉中锌水平明显升高,说明补锌可以提高冷暴露大鼠的组织锌水平(骆文静等,2004)。关于锌可以提高大鼠耐寒力的机制目前还不确定。有学者认为由锌组成的许多酶的活性基团可以抑制自由基的产生,保护细胞膜和蛋白免受自由基的氧化损伤,从而提高机体的耐寒力。
5.3.4维生素
妊娠后期补充维生素E可能会通过提高胎儿BAT的量,而影响胎儿的能量状况(Nockels等,1996;Thomas等,1995)。苗振春等(2001)报道了全反式维甲酸(tRA)对棕色脂肪组织的影响,tRA能使棕色脂肪组织线粒体中UCP含量增加,但不改变其活性;能增加棕色脂肪组织中脂肪消耗,可能在白色脂肪组织中诱导产生UCP。Puigserver等(1996)发现用tRA处理大鼠,其肩胛间BAT体积明显变小,BAT中总蛋白含量减少,但特异性UCP却增加2倍,说明tRA能引起UCP含量增加,同时增加脂肪消耗,但其确切机制仍不明确,有待进一步研究。
5.3.5母体营养的影响
孕期1~2周,母老鼠营养不良,子宫内营养不良,使雄性子鼠增重和脂肪沉积下降,尽管正常采食,但血浆中TG浓度下降;使雌性子鼠BAT重量和蛋白质含量下降,细胞色素C氧化酶活力下降,表现为低交感神经活动(Anguita,1993)。Luz等(2003)报道给孕期母鼠限食(50%),子鼠出生后耐寒冷的能力差。由于子宫内营养不良,可能推迟下丘脑-甲状腺轴的发育,反过来影响BAT的发育,导致新生儿产热不足。
5.3.6激素和转录因子
结语
6结语
动物与环境之间的关系一直是动物学家们所关心的问题,动物只有与环境条件相协调相适应才能够顺利生存下去。动物对环境的适应是一个能动的过程,动物通过启动与抗冷性有关的基因表达,给出在逆境中的保护措施,才能协助动物自身渡过难关。生物机体形成冷适应后,表现出基础代谢或非寒颤性产热增加。对棕色脂肪组织产热机理和代谢的研究,可以扩大动物生态位和动物对环境骤然变化的适应能力,对提高幼畜的成活率十分重要。BAT中的UCP1和PGC-1α可调节能量平衡及底物代谢,影响肥胖的发生,选择性提高UCP1和PGC-1α活性是减肥的有效方法,可为肥胖的治疗研究提供理论依据。总之,弄清BAT的功能对发展畜牧业和人类健康都具有重要的现实意义。
最新研究
2022年6月29日消息,哈佛医学院乔斯林糖尿病中心研究团队发现,棕色脂肪组织在寒冷刺激时产生的一种天然分子,可以消解暴露在低温环境中肥胖引发的慢性炎症,同时改善饮食诱导的肥胖小鼠胰岛素敏感性和葡萄糖代谢。
参考资料
[1]
科学新发现丨科学家发现寒冷能减肥的关键机制 · 中国科技网[引用日期2022-06-29]
棕色脂肪组织相关的文章

脑外伤多由头部突然的加速或减速运动、头部快速撞击不能移动的硬物、高空坠落、跌倒、头部遭受暴力等造成的脑损伤。交通事故伤是脑外伤最常见的原因,其次为坠落伤、火器、暴力、摔倒、异物贯通、自然灾害、工伤事故等。脑外伤不同的损伤部位症状不同,包括运动、感觉、言语、视觉、听觉等方面异常。脑外伤急性期时治疗以维
本雅明·内塔尼亚胡(בִּנְיָמִין נְתַנְיָהוּ,Benjamin Netanyahu),男,1949年10月21日出生于以色列特拉维夫-雅法,毕业于麻省理工学院政治家、军人,第九、十三任以色列总理,兼任外交部长、国防部长、卫生部长、移民部长。
亚得里亚海,意大利语作Mare Adriatico,英语作Adriatic sea。地中海的一个大海湾。长约800公里(500哩),平均宽160公里(100哩),平均水深444公尺(1,457呎),最深处1,324公尺(4,035呎),面积131,050平方公里(50,590平方哩)。北浅、南深,平

尚可名片
这家伙太懒了,什么都没写!
作者
